

Глава 7. Частная физиология центральной нервной системы
Частная физиология ЦНС изучает функции структур головного и спинного мозга, а также механизмы их осуществления.
7.1. Нервные центры и методы их исследования
Понятие о нервном центре. Нервный центр — это совокупность нервных клеток, регулирующих определенную функцию организма. Эти нервные клетки могут быть расположены компактно в пределах одной анатомической структуры или представлять собой функционально объединенные группы нейронов, располагающиеся во многих отделах центральной нервной системы. Например, нервный центр, регулирующий дыхание, имеет свое представительство в продолговатом и спинном мозге, в гипоталамусе, лимбической системе и коре мозга.
Термин "нервный центр" применяется также д ля обозначения места замыкания рефлекторных реакций. В этом случае называют отдел ЦНС или конкретное ее образование, в котором происходит переключение возбуждения с афферентного на эфферентное звено рефлекторной дуги. Например, нервным центром замыкания коленного рефлекса являются 2— 3-й поясничные сегменты спинного мозга.
Методы исследования функций нервных центров. Выделяют следующие методы:
1) электроэнцефалография методика регистрации биопотенциалов, генерируемых головным мозгом, при отведении их от поверхности кожи головы. Величина таких биопотенциалов составляет 1—300 мкВ. Они отводятся с помощью электродов, налагаемых на поверхность кожи во всех областях головы. Затем с использованием прибора электроэнцефалографа регистрируется электроэнцефалограмма (ЭЭГ) графическая кривая, отражающая непрерывные изменения
Р-ритм
/\M/VWWWWWWWW\MAAA/WWWW\ — ot-ритм
л4А!М1ШЛ1\ЛЛЛ^
о-ритм
t
t
Бодрствование
Включен свет
в
темноте реакция десинхронизации
б
а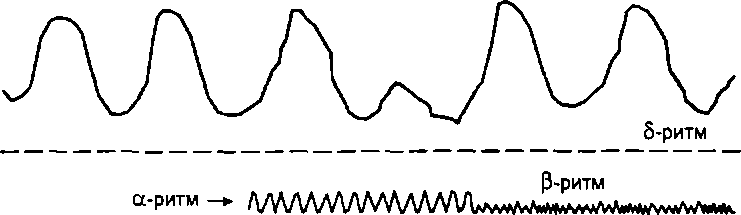
Рис. 7.1. Регистрация биопотенциалов головного мозга при отведении
от поверхности головы: а - основные ритмы электроэнцефалограммы (схема); б — смена а-ритма на Р-ритм (реакция десинхронизации) при активации ретикулярной формации под действием света
(волны) биопотенциалов мозга. Частота и амплитуда электроэнцефалографических волн отражают уровень активности нервных центров. Выделяют четыре основных ритма ЭЭГ(рис. 7.1).
Альфа-ритм имеет частоту 8-13 Гц и амплитуду 30- 70 мкВ. Это ритм бодрствующего покоя. Он выявляется приблизительно у 90% людей, когда они находятся в покое с закрытыми глазами или в темноте. Этот ритм наиболее выражен в затылочных областях коры.
Бета-ритм характеризуется частотой 14—30 Гц и амплитудой до 30 мкВ. Это мелкие нерегулярные волны. Данный ритм особенно выражен в лобных и апикальных областях коры. Он свидетельствует об активном, деятельном состоянии мозга. Если другие низкочастотные ритмы сменяются на бета- ритм, то это свидетельствует о переходе мозга к большей функциональной активности. Такую реакцию называют реакцией десинхронизации или активации.
Тета-ритм имеет частоту 4-7 Гц и амплитуду до 200 мкВ. Он проявляется при поздних стадиях засыпания человека и развитии наркоза. Может возникать и в бодрствующем состоянии при длительном эмоциональном напряжении.
Дельта-ритм имеет частоту 0,5—3 Гц и большую (до 300 мкВ) амплитуду волн. Он устанавливается по всей поверхности мозга во время глубокого сна или наркоза.
Считается, что водителями ЭЭГ-ритмов могут быть тала- мус и ствол мозга. Причем, таламус индуцирует и навязывает коре высокочастотные, а ствол мозга — низкочастотные (тета и дельта) ритмы;
метод вызванных потенциалов позволяет регистрировать изменение электрических потенциалов коры в ответ на стимуляцию различных рецепторных полей или проводящих путей. Возникающие в ответ на одномоментное раздражение биопотенциалы коры носят волнообразный характер, длятся до 300 мс. Для выделения вызванных потенциалов из спонтанных электроэнцефалогических волн применяют сложную компьютерную обработку ЭЭГ Эта методика используется в эксперименте и в клинике для определения состояния рецептор- ной, проводниковой и центральной частей анализаторов;
микроэлектродный метод позволяет с помощью тончайших электродов, вводимых в клетку или располагаемых в ее ближайшем окружении, регистрировать электрическую активность отдельных нейронов, а также раздражать их;
стереотаксический метод позволяет вводить в заданные структуры мозга иглы, зонды, электроды с лечебной и диагностической целью. В настоящее время разработаны'сте- реотаксические атласы, в которых указывается, под каким углом и на какую глубину относительно характерных ориентиров черепа вводится игла или электрод. При этом голова больного фиксируется в специальном держателе;
метод раздражения различных структур мозга чаще всего осуществляется с помощью слабого электрического тока. Такое раздражение легко дозируется, не вызывает повреждений нервных клеток и может наноситься многократно. В качестве раздражителей используются также различные биологически активные вещества;
методы перерезок, экстирпации и функциональной блокады нервных центров широко использовались в эксперименте в начальный период накопления знаний о мозге.
Но и в наше время клинические наблюдения за изменением регуляции функций у больных, подвергшихся удалению мозговых структур (при опухолях, кровоизлияниях, травмах), дают все новые сведения о физиологической роли этих структур.
При функциональной блокаде производят временное выключение функций нервных центров путем введения биологически активных веществ, воздействий специальными электрическими токами, охлаждения, ультразвука, рентгеновских лучей;
реоэнцефалография — методика исследования пульсовых изменений кровенаполнения мозговых сосудов. Она основана на измерении сопротивления тканей головы электрическому току;
эхоэнцефалография позволяет определять локализацию и размеры образований и полостей мозга и черепной коробки. Эта методика основывается на регистрации ультразвуковых волн, отраженных от тканей головы;
методы компьютерной томографии, основанные на регистрации ядерного магнитного резонанса тканей головы и прохождения через них рентгеновских лучей, дают четкие послойные изображения структур мозга;
методы условных рефлексов и регистрации вызванных и спонтанных поведенческих реакций позволяют исследовать интегративные функции высших отделов мозга. Эти методы подробнее рассмотрены ниже.
7.2. Спинной мозг
Для изучения физиологии спинного мозга необходимо напомнить важнейшие сведения по его анатомической структуре. Спинной мозг с его оболочками располагается в позвоночном канале от уровня большого затылочного отверстия до 2-го поясничного позвонка. Спинной мозг подразделяется на сегменты: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый. Сегментом называют участок, от которого симметрично отходят левая и правая пары спинномозговых корешков. В каждой паре есть так называемый задний (дорсальный) и передний (вентральный) корешок.
Через задние корешки в спинной мозг входят афферентные (центростремительные, чувствительные) волокна. Они являются отростками нейронов, тела которых располагаются в спинальных ганглиях. Передние корешки начинаются от вентральной поверхности сегментов спинного мозга. Они образованы аксонами эфферентных (центробежных, двигательных) нейронов, тела которых находятся в передних рогах спинного мозга, а также отростками вегетативных нейронов, тела которых расположены в боковых рогах. Перед выходом из межпозвоночных отверстий передний и задний корешки объединяются в спинномозговой нерв, который идет к строго определенным сегментам кожи, мышцам и внутренним органам.
Функции, центры и нейроны спинного мозга. Важнейшие функции спинного мозга: рефлекторная (замыкательная), проводниковая и интегративная. Эти функции осуществляются благодаря спинальным нейронам и нервным волокнам, формирующим проводящие пути.
Нейроны спинного мозга. Расположены в тяжах серого вещества, которое в совокупности с промежуточным веществом располагается вокруг центрального спинномозгового канала, заполненного цереброспинальной жидкостью.
Поперечный срез серого вещества по форме напоминает крылья бабочки. В нем выделяют передние и задние рога, содержащие нейроны, различающиеся по морфологии и функциям. В грудных и двух верхних поясничных сегментах имеются также боковые рога (выступы) серого вещества. В них располагаются вегетативные нейроны, эфферентные волокна от которых идут к вегетативным ганглиям. В задних рогах сосредоточены тела вставочных нейронов, выполняющих замыкатель- ную функцию. Эти нейроны передают нервные импульсы от афферентных к эфферентным нейронам в пределах одного спинального сегмента. Здесь же имеются многочисленные ассоциативные нейроны, передающие импульсацию от афферентных нейронов к выше и ниже расположенным сегментам спинного мозга.
Эфферентные нейроны спинного мозга представлены а- и у-мотонейронами, расположенными в передних рогах серого вещества, и вегетативными нейронами, находящимися в боковых рогах. Большинство а-мотонейронов имеют крупные тела. Максимальная частота потенциалов действия, которую они могут генерировать, составляет лишь 50 импульсов в секунду. Это вызвано тем, что потенциал действия а-мотонейронов имеет длительную следовую гиперполяризацию (до 150 с), во время которой возбудимость клетки снижена. Кроме того, имеется еще специальный тормозной механизм в виде простейшей нейронной цепи: а-мотонейрон клетка Рению. Благодаря ветвлению аксона мотонейрона каждый проходящий по нему потенциал действия активирует клетку Реншо, а она тормозит мотонейрон.
Центры спинного мозга. В спинном мозге находятся центры, участвующие в регуляции большинства функций, органов и систем организма.
Говоря о центрах спинного мозга, имеют в виду сегменты, в которых замыкаются спинальные рефлексы и отделы спинного мозга, где сосредоточены нейронные группы, обеспечивающие регуляцию определенных физиологических актов и реакций. Например, спинальные жизненно важные отделы дыхательного центра представлены мотонейронами передних рогов 3—5-го шейных и средних грудных сегментов. Если эти отделы мозга повреждены, то наступает смерть от остановки дыхания.
Зоны
корешков
Рис.
7.2.
Метамерия в афферентной иннервации
кожи (области распространения афферентных
волокон корешков спинного мозга)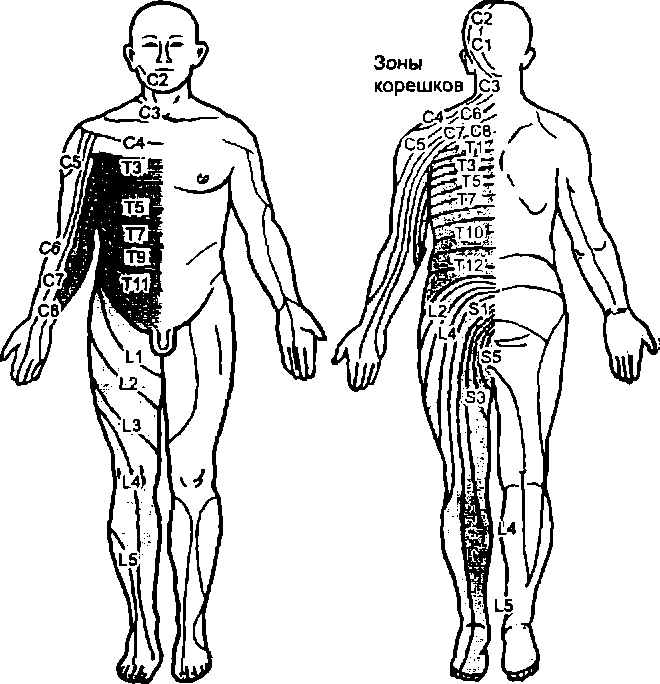
В иннервации скелетных мышц, гладких мышц внутренних органов и особенно структур кожи тела выявляется метамерия (рис. 7.2). Мышцы шеи управляются моторными центрами шейных слинальных сегментов CI — С4, диафрагмы — сегментами СЗ—С5, рук — С5—Th2, туловища — Th3—L1, ног — L2— S5. Кожа шеи и рук иннервируется из верхних (шейных) сегментов спинного мозга, область туловища — грудных, ног — поясничных и крестцовых сегментов.
Области распространения окончаний нервных волокон, идущих от соседних спинальных сегментов к структурам тела, частично перекрываются: каждый сегмент иннервирует не только свой метамер, но и половину выше и ниже лежащего метамера. Таким образом, каждый метамер тела получает иннервацию от трех сегментов спинного мозга, а волокна одного сегмента имеют свои синаптические окончания в трех метамерах.
Замыкательная функция спинного мозга. Рефлексы, замыкание рефлекторной дуги которых происходит в спинном мозге, называют спинальными. Спинальные рефлексы подразделяют по их функциональной значимости и особенностям рефлекторной дуги. Выделяют миотатические, оборонительные сгибательные, перекрестные разгибательные и вегетативные спинальные рефлексы.
• Миотатические рефлексы. Рефлексы, возникающие в ответ на раздражение рецепторов мышечных веретен, называют миотатическими, сухожильными или рефлексами растяжения.
Мышечные веретена представляют собой вытянутые структуры, имеющие в центре соединительнотканную капсулу (ядерную сумку), в которой располагаются нервные окончания. С каждой стороны к ядерной сумке прикреплены миниатюрные (интрафузальные) мышечные волокна. Их сокращение настолько мало, что не вызывает видимого укорочения целостной скелетной мышцы, но оно достаточно для растяжения ядерной сумки и активации находящихся там рецепторов растяжения. Мышечные веретена располагаются параллельно обычным (экстрафу- зальным) волокнам скелетной мышцы.
Адекватным раздражителем для рецепторов мышечных веретен является растяжение. Эти рецепторы активируются при: 1) растяжении всей мышцы; 2) сокращении миниатюрных интрафузальных волокон, входящих в структуру веретен. Сокращение интрафузальных волокон запускается импульсами, Приходящими по аксонам у-мотонейронов спинного мозга.
Импульсация от рецепторов растяжения мышечных веретен передается в спинной мозг по быстропроводящим афферентным волокнам (Аа), которые проникают непосредственно в передние рога спинного мозга и передают возбуждение непосредственно на а-мотонейроны. Таким образом, миотатические рефлексы, вызываемые с рецепторов растяжения мышечных веретен, имеют двухнейронную рефлекторную дугу и только один центральный синапс (рис. 7.3). Такие рефлексы называют моносинаптическими. Они замыкаются в том же спинальном сегменте, в который входит афферентное волокно. Все другие рефлексы, замыкающиеся в центральной нервной системе, —полисинаптические.
Миотатические рефлексы имеют значение в регуляции тонуса и степени сокращения скелетных мышц. Во врачебной практике эти рефлексы используются для тестирования степени возбудимости спинальных моторных центров, а также для диагностики нарушения проведения возбуждения в рефлекторной дуге и определения уровня повреждения спинного мозга.
Рис.
7.3.
Пути замыкания некоторых спинальных
рефлексов: а
— схема простейших рефлекторных дуг:
1) с рецепторов мышечных веретен; 2) с
сухожильных рецепторов Гольджи; б —
схема простейшей рефлекторной дуги
оборонительного сгибательного рефлекса,
вызываемого раздражением кожи
(одновременно с сокращением сгибателя
происходит расслабление разгибателя)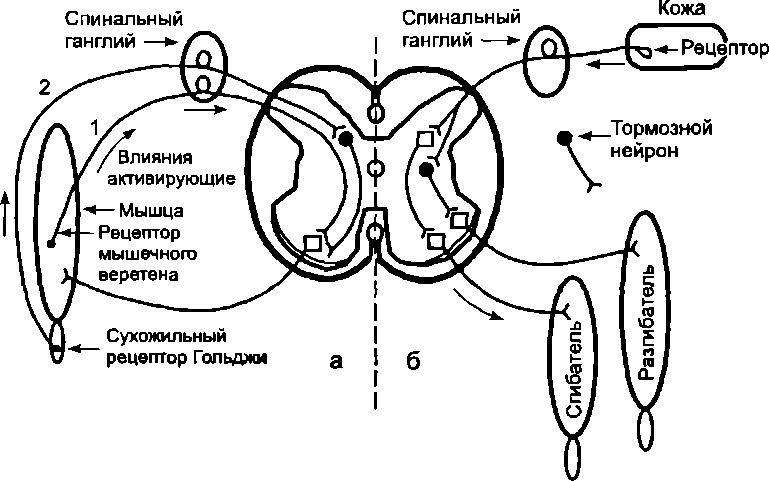
Для вызова сухожильного рефлекса производят удар неврологическим молоточком по сухожилию той мышцы, сокращение которой хотят вызвать. При этом происходит кратковременное растяжение мышцы и активация рецепторов мышечных веретен. Например, для вызова ахиллова рефлекса производят удар по ахиллову сухожилию. В ответ на это воздействие через доли секунды происходит подошвенное сгибание стопы. Хотя для таких рефлексов часто применяют название сухожильные, следует понимать, что запуск этих рефлексов происходит с рецепторов той мышцы, с которой связано сухожилие (ахиллов рефлекс запускается с рецепторов икроножной мышцы, коленный — с четырехглавой мышцы бедра).
Центр замыкания ахиллова рефлекса находится в 1—2-м крестцовом сегменте спинного мозга, коленного рефлекса — во 2—3-м поясничном сегменте, рефлекса сгибания предплечья — в 4—6-м шейном сегменте.
В самих сухожилиях также имеются рецепторы. Они образуются окончаниями афферентных волокон, обвивающих группы сухожильных нитей (рецепторы Гольджи). Эти рецепторы активируются при сильном натяжении сухожилий. Импульсация от них тормозит активность спинальных мотонейронов, препятствуя тем самым чрезмерному напряжению мышцы, связанной с сухожилием, и разрыву сухожилия. Благодаря функциям сухожильных рецепторов регулируется прежде всего напряжение мышц, а благодаря рецепторам мышечных веретен — длина мышц. Рефлексы, вызываемые с сухожильных рецепторов Гольджи, являются полисинаптическими, оказывают влияние на мотонейроны через вставочный тормозной нейрон.
Оборонительные сгибательные рефлексы. Эти рефлексы возникают в ответ на действие повреждающих или новых неожиданных раздражителей на самые разные рецепторы конечностей и туловища (за исключением рецепторов ядерной сумки мышечных веретен) и проявляются сгибанием конечностей. Таким образом достигается устранение контакта тела с повреждающим или потенциально опасным агентом. Например, укол кожи руки вызывает сгибание руки еще до того, как человек подумал об этом движении и приложил волевое усилие.
Перекрестные разгибательные рефлексы. Выше мы Рассмотрели упрощенную схему оборонительной реакции сгибательного рефлекса. Фактически же для достижения до- сточной эффективности сгибания конечности необходимо,
чтобы наряду с сокращением сгибателей происходило расслабление ее разгибателей. Это и достигается за счет перекрестного разгибательного рефлекса (рис. 7.3).
Одновременное и противоположно направленное изменение активности сгибателей и разгибателей называют перекрестным рефлексом. Эти рефлексы возникают в ответ на действие тех же раздражителей, которые вызывают сгиба- тельные рефлексы. Они способствуют устранению контакта тела с повреждающими агентами. Рефлекторная дуга перекрестных разгибательных рефлексов формируется одновременно с осуществлением сгибательного рефлекса за счет механизма реципрокных взаимодействий нервных центров.
При реципрокном взаимодействии возбуждение одного нервного центра за счет жестко запрограммированных коротких цепочек нейронов тормозит активность другого. Такие взаимодействия моторных центров мышц-антагонистов в спинном мозге обеспечивают адекватные двигательные реакции конечностей. Сгибательные и перекрестные разгибательные рефлексы являются составляющими общей двигательной реакции.
Мы рассмотрели лишь ответную реакцию мышц одной конечности. Но при сильных воздействиях, как правило, происходят содружественные сокращения мышц обеих конечностей. В координации этих сокращений имеют значение реципрокные взаимоотношения моторных центров одноименных мышц противоположных конечностей. Фактически в оборонительной реакции на действие раздражителя можно выделить четыре составляющие.
Так, при нанесении на кожу левой ноги болевого воздействия будет рефлекторно возбуждаться сгибатель этой ноги и одновременно тормозиться ее разгибатель. В то же время будет тормозиться сгибатель правой ноги и возбуждаться ее разгибатель. Такие взаимодействия спинальных двигательных центров являются основой для возникновения ритмических двигательных реакций на раздражение. Эти взаимодействия являются также одним из показателей наличия интегративных процессов в спинном мозге.
• Вегетативные рефлексы. Спинальные вегетативные рефлексы участвуют в регуляции функций внутренних органов, кровообращения, обмена веществ и секреции желез.
Структура и механизмы реализации вегетативных рефлексов подробно рассматриваются в § 7.11 (физиология автономной нервной системы).
Интегративная функция спинного мозга
В спинном мозге имеется много нейронов (проприоспинальных), которые обеспечивают связи между сегментами, расположенными на всех уровнях. Эти нейроны участвуют в более сложных регуляциях, в которых обнаруживается интегративная функция спинного мозга.
Одним из проявлений интегративной функции является способность спинного мозга управлять ритмическими, длительно проявляющимися, координированными двигательными реакциями, в которых задействованы мышцы разных конечностей и туловища. Эти реакции осуществляются и при потере связи спинного мозга с головным. Такая функция спинного мозга осуществляется благодаря наличию и функционированию специализированных нейронных цепей, в которых запрограммированы некоторые ритмические лакомоторные акты, обеспечивающие перемещение тела в пространстве. После вызова пусковым стимулом такие реакции могут поддерживаться некоторое время автоматически.
Особенно четко интегративная функция спинальных нейронов может быть прослежена на примере регуляции тонуса мышц туловища и конечностей. Тонус мышц поддерживается за счет тонуса моторных центров спинного мозга (т.е. за счет наличия постоянной импульсации от а-мотонейронов этих центров к скелетным мышцам). Тонус а-мотонейронов поддерживается благодаря интеграции импульсов, приходящих к этим нейронам от многочисленных источников: рецепторов, расположенных в самих мышцах, коже, внутренних органах, и от ряда двигательных центров головного мозга (рис. 7.4). Далее будут рассмотрены нисходящие пути, передающие импульсации к спинальным мотонейронам. Мотонейрон выступает интегратором этих сигнализаций и под их влиянием генерирует определенную частоту импульсов, посылаемых к мышцам.
У взрослого человека интегративная функция спинного мозга проявляется чаще всего как составляющая в регулятор- ных реакциях, осуществляемых целостной нервной системой и обеспечивающих приспособительные поведенческие акты и 0егетативные регуляции.
Степень интегрирования функций спинного мозга с функциями головного мозга такова, что при потере связей с головам мозгом возникают явления спинального шока.
Нисходящие
пути от
коры и ядер ствола
мозга
Рис.
7.4.
Интеграция на а-мотонейронах импульсаций
от сенсорных рецепторов и нисходящих
путей от коры и ствола мозга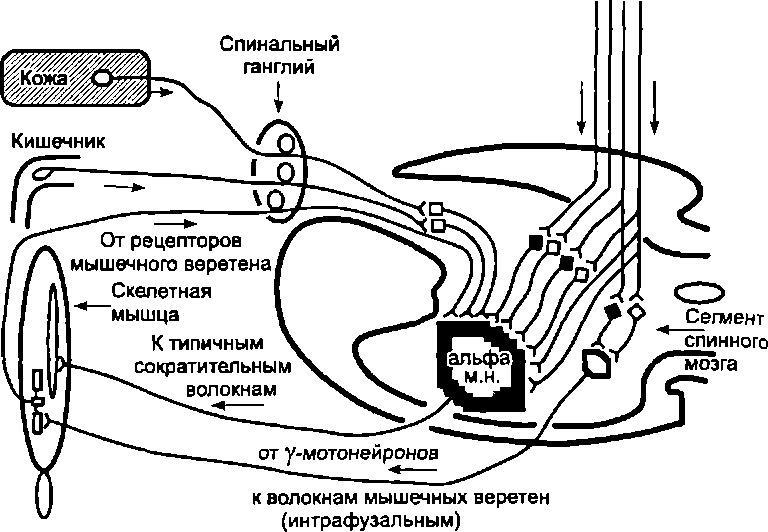
Спинальный шок возникает при разрыве спинного мозга и проявляется резким снижением возбудимости и угнетением функций нервных центров, расположенных ниже места повреждения. При этом исчезает большинство рефлексов, регулирующих движения и тонус скелетных мышц, нарушаются вегетативные функции, акты мочеиспускания и дефекации. Кровяное давление падает, но относительно раньше других вегетативных функций начинает повышаться вместе с восстановлением ряда рефлексов, регулирующих перераспределение крови между сосудистыми областями. Если повреждение произошло выше 3-го шейного сегмента, то наступает остановка дыхания и смерть, если ниже шейных сегментов - дыхание может сохраниться за счет сокращений диафрагмы. Современная медицина дает возможность спасти жизнь таким больным.
Проводниковая функция спинного мозга. Спинной мозг проводит нервные импульсы в восходящем направлении к головному мозгу и в нисходящем — от головного мозга к спи- нальным сегментам. Значительную часть проводящих путей сПинного мозга составляют аксоны так называемых проприо- спинальных нейронов. Волокна этих нейронов создают связи между спинальными сегментами и не выходят за пределы спинного мозга.
Восходящие (чувствительные) пути образуются волок- нами, являющимися отростками: 1) афферентных нейронов, тела которых располагаются в спинальных ганглиях; 2) ассоциативных нейронов, тела которых находятся в задних рогах спинного мозга, а аксоны идут к структурам ствола мозга таламуса и мозжечка.
Ход волокон, передающих сигнализацию разной чувствительности (модальности), неодинаков. Проводящие волокна от проприорецепторов передают к головному мозгу сигнализацию о состоянии мышц, сухожилий, суставов. Эти волокна являются отростками нейронов спинальных ганглиев. Войдя через задние корешки в спинной мозг, они по той же стороне спинного мозга (не делая перекреста), в составе тонкого и клиновидного пучков восходят до нейронов продолговатого мозга, где передают импульсацию на второй афферентный нейрон (рис. 7.5). Этот нейрон проводит импульсацию к ядрам таламуса. После переключения на нейронах таламуса импульсация доходит до коры мозга и обеспечивает возникновение ощущения степени напряжения мышц, положения конечностей и угла сгиба в суставах, пассивного движения, вибрации.
В составе тонкого пучка также проходит часть волокон от рецепторов кожи, обеспечивающих тактильную чувствительность: прикосновение, давление, вибрацию.
Остальные спинальные чувствительные пути образованы отростками вторичных афферентных нейронов, тела которых находятся в задних рогах спинного мозга. Аксоны этих нейронов делают перекрест и по противоположной стороне спинного мозга в составе латерального спиноталамического тракта идут к таламусу. В этом тракте расположены, в частности, волокна, обеспечивающие болевую и температурную чувствительность, а также часть волокон тактильной чувствительности (рис. 7.5).
В боковых канатиках также находятся передний и задний спиномозжечковые тракты. Они передают импульсацию от Проприорецепторов к мозжечку. Здесь же проходят пути от спинальных сегментов к покрышке, оливе и ретикулярной формации ствола мозга.
Рис.
7.5.
Ход проводящих путей от проприорецепторов,
тактильны*, температурных и болевых
рецепторов к стволу и коре мозга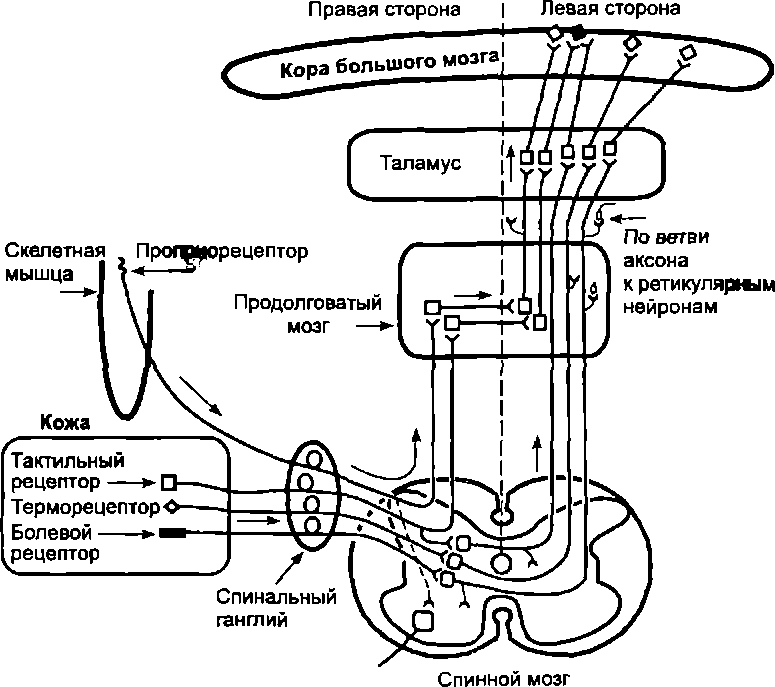
Нисходящие (двигательные) проводящие пути, регулирующие произвольные движения, представлены двумя корково- спинальными трактами. Латеральный корково-спинальный путь идет в боковых канатиках белого вещества мозга, а передний корково-спинальный путь — в передних канатиках. Эти пути называют также пирамидными. Они проходят от пирамидных нейронов моторной зоны коры больших полушарий к передним рогам спинного мозга и передают импульсацию (как прямо, так и через вставочные нейроны) на мотонейроны. Эта импульсация регулирует произвольные движения человека.
Имеются также двигательные пути, исходящие не от коры, а от ствола мозга. Эти пути часто называют экстрапирамидными. В боковых канатиках находятся красноядерно-спинномоз- говой и оливо-спинномозговой пути. В передних канатиках располагаются преддверно-спинномозговой и ретикулярно-
спинномозговой пути. По этим путям к спинальным мотонейронам передаются импульсы, обеспечивающие непроизвольные двигательные акты, совершающиеся за счет врожденных или приобретенных рефлексов и двигательных программ.
Считается также, что эти пути могут быть использованы и для осуществления произвольных двигательных реакций, благодаря тому, что кора мозга может управлять функциями экстрапирамидной системы через специальные (кортикофу- гальные) нисходящие тракты.
7.3. Продолговатый мозг
Продолговатый мозг является частью головного мозга, соединяющей его со спинным мозгом. Его строение во многом сходно со строением спинного мозга. Центральный канал спинного мозга, заполненный цереброспинальной жидкостью, на уровне продолговатого мозга переходит в полость четвертого желудочка головного мозга. На вентральной поверхности четвертого желудочка располагается ромбовидная ямка, являющаяся ориентиром месторасположения ряда жизненно важных нервных центров (рис. 7.6).
Продолговатый мозг выполняет функции: рефлекторную, проводниковую и интегративную.
Рефлекторная функция продолговатого мозга. В продолговатом мозге замыкается ряд защитных рефлексов: мигательный, слезоотделительный, рвотный, чихательный, кашлевой, а также рефлексы, регулирующие тонус мышц и положение тела в пространстве. Для их осуществления, как правило, требуется участие нейронов моста. Эти рефлексы также называют установочными и позотоническими. Они обеспечивают распределение тонуса разных мышечных групп, необходимого для поддержания правильной позы тела в покое и готовности к выполнению рабочих движений.
161
®Зак. 181
Рис.
7.6.
Схема представительства ядер черепных
нервов в
стволе мозга (ядра III—XII
черепных нервов)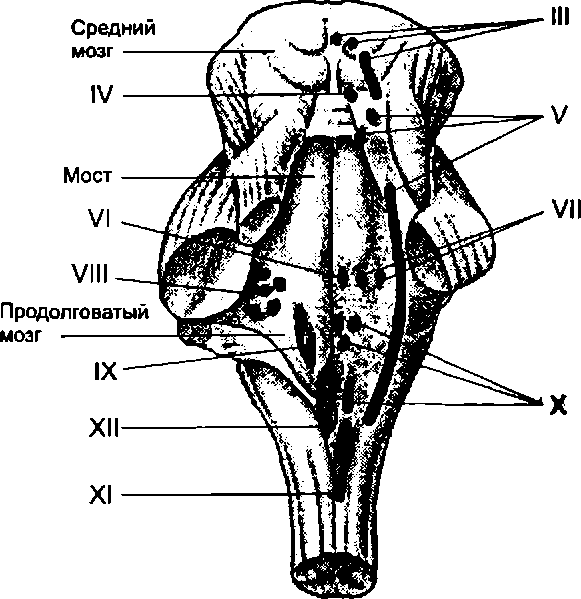
ложение головы в пространстве относительно направления силы тяжести (гравитационного поля земли).
В продолговатом мозге замыкается также ряд рефлексов, регулирующих вегетативные функции и гомеостаз. Они рассматриваются в разделах физиологии дыхания, кровообращения, пищеварения и терморегуляции.
В осуществлении вышеназванных рефлекторных реакций важна роль IX—XII пар черепных нервов, имеющих ядра в продолговатом мозге (рис. 7.6).
Проводниковая и интегративная функции продолговатого мозга. Через продолговатый мозг проходят чувствительные проводящие пути от спинного мозга к таламусу, мозжечку и ядрам ствола. Расположение этих путей в белом веществе сходно с таковым в спинном мозге. В дорсальном отделе продолговатого мозга находятся тонкие и клиновидные ядра, на нейронах которых заканчиваются одноименные пучки афферентных волокон, идущих от мышц, суставов и тактильных рецепторов кожи. Импульсация от таких волокон переключается на вторичные афферентные нейроны тонкого и клиновидного ядер.
В латеральной области белого вещества проходят нисходящие двигательные пути: оливо-спинальный и рубро-спиналь- ный. В вентральной части находится корково-спинальный двигательный тракт. Его волокна на уровне продолговатого мозга сгруппированы в образования, называемые пирамидами. Большинство (до 80%) этих волокон переходит на противоположную сторону, формируя перекрест пирамид. Остальные (до 20%) неперекрещенных волокон переходят на противоположную сторону уже на уровне спинного мозга.
По черепным нервам в продолговатый мозг вступают также волокна, проводящие импульсы от рецепторов мышц и кожи лица, дыхательных путей и рта, интерорецепторов пищеварительной и сердечно-сосудистой систем.
Интегративная функция продолговатого мозга проявляется в реакциях, которые не могут быть отнесены к простым рефлексам. В его нейронах запрограммированы некоторые сложные регуляции, требующие для своего осуществления участия центров других отделов нервной системы, взаимодействия с ними. Некоторые ретикулярные нейроны обладают ав- томатией, тонизируют и координируют активность нервных центров.
Центры продолговатого мозга. В продолговатом мозге находится ряд необходимых для сохранения жизнедеятельности нервных центров. Среди них центры регуляции дыхания, кровообращения, ряда пищеварителных функций. Разрушение или блокада жизненно важного отдела дыхательного центра ведет к немедленной остановке дыхания и смерти. Подробнее структура и функция жизненно важных центров продолговатого мозга рассмотрена в разделах частной физиологии.
В продолговатом мозге расположены также ядра IX— XII пар черепных нервов (рис. 7.6).
Центры языкоглоточного нерва (IX пара) включают чувствительное, двигательное и парасимпатическое ядра. Эти ядра воспринимают импульсацию от вкусовых, тактильных, болевых и температурных рецепторов слизистой оболочки глотки, части языка, интерорецепторов каротидного тельца. Они участвуют в рефлексах жевания, глотания, регуляции пищеварения и кровообращения. Эфферентная импульсация от парасимпатического ядра стимулирует секрецию околоушной слюнной железы.
Расположенные в продолговатом мозге ядра блуждающего нерва (X пара) являются источником чувствительной иннервации неба, корня языка, органов шеи, груди и брюшной полости. Заднее ядро обеспечивает парасимпатическую иннервацию сердца и дыхательной системы, гладкомышечной и железистой ткани области лица, шеи, органов грудной и брюшной полостей. Это ядро участвует в рефлекторных регуляциях функций названных органов.
Двойное двигательное ядро включается в эфферентное звено рефлексов глотания, кашля, чихания, рвоты. Оно участвует также в проведении импульсаций к мышцам гортани, регулирующим высоту и тембр голоса.
Ядро добавочного нерва (XI пара) является источником эфферентной иннервации грудино-ключично-сосцевидной и трапециевидной мышц, сокращения которых вовлекаются в осуществление наклонов головы, поднимания плечевого пояса и смещения лопаток.
Ядра подъязычного нерва (XII пара) располагаются в области нижней части ромбовидной ямки и трех верхних сегментов спинного мозга. Этот нерв содержит двигательные волокна, регулирующие сокращения мышц языка, участвующие в обеспечении приема и обработки пищи, а также осуществлении речи.
На границе продолговатого мозга и моста расположены чувствительные ядра преддверно-улиткового нерва (VIII пара). К ним приходит импульсация от органов слуха и равновесия. Здесь она переключается на вторые афферентные нейроны, которые проводят информацию к таламусу и среднему мозгу, участвуют в механизмах замыкания ориентировочного рефлекса, статических, статокинетических, вестибулоглазных и вестибуловегетативных рефлексов.
7.4. Мост
Мост расположен между продолговатым и средним мозгом (рис. 7.6). Его проводящие пути и функции во многом схожи и интегрированы с продолговатым мозгом, поэтому эти два участка ствола мозга иногда рассматривают как единую структуру.
В дорсальном отделе моста располагаются чувствительные пути, идущие к таламусу, а в вентральной части — двигательные пути от коры и красного ядра. Мост является также связующим звеном по обмену информацией между мозжечком и другими отделами ЦНС. На ядрах моста заканчиваются волокна корково-мостового двигательного пути. Импульсация, идущая от коры, после переработки на нейронах ядер моста передается к мозжечку по аксонам этих нейронов, проходящих в составе средних ножек мозжечка.
В ретикулярной формации моста расположены ядра, формирующие медиальный ретикулоспинальный тракт. Волокна этого тракта без перекреста идут к спинальным а- и у- мотонейронам. Передаваемая импульсация возбуждает мотонейроны разгибателей и тормозит нейроны сгибателей.
В дорсальной части серого вещества моста располагаются ядра V—VIII пары черепно-мозговых нервов. Эти центры обеспечивают чувствительность кожи лица и головы, вкусовую чувствительность, слух и восприятие изменений положения головы.
В ретикулярной формации моста находится один из отделов дыхательного центра (пневмотаксический отдел). Он участвует в формировании частоты и ритма дыхательных движений.
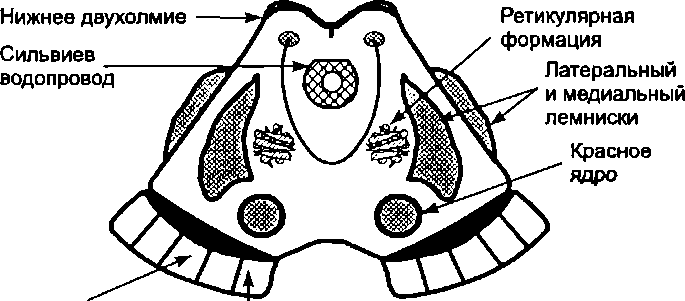
7.5. Средний мозг
Средний мозг располагается между мостом и промежуточным мозгом (рис. 7.7, 7.8). Его подразделяют на область ножек мозга и крышу мозга. Через средний мозг проходит узкий
Кортикоспинапьный
тракт Кортикомостовой
тракт
Рис.
7.7.
Схема поперечного разреза среднего
мозга
Краснов;"
«жаг» х
..ядро / V.
"
' ^чА^СХ Гипоталамус
'Средний^"4
.мозг
в|5 I
Мозжечок
Мост
Ретикулярная формация (мостовая часть)
' Вестибулярные ядра Продолговатый ^Ретикулярная формация моэг
' Спинной мозг
Рис. 7.8. Схема расположения некоторых ядер ствола мозга
и гипоталамуса. Ядра гипоталамуса: I — паравентрикулярное,2 — дорсомедиальное,3 — преоптическое, 4 — супраоптическое,5 — заднее
канал — водопровод мозга, заполненный ликвором и соединяющий третий и четвертый желудочки.
Ножки мозга занимают вентральную часть среднего мозга, крыша — дорсальную. На крыше среднего мозга выступают два парных образования: верхние и нижние холмики (бугорки).
В сером веществе покрышки среднего мозга находятся ядра глазодвигательного (III пара) и блокового (IV пара) черепных нервов (соответственно на уровне верхних и нижних холмиков). Латерально от водопровода расположена ретикулярная формация и ядро среднемозгового пути тройничного нерва. Кроме того, в состав среднего мозга входят красное ядро и черная субстанция, а также восходящие и нисходящие проводящие пути, располагающиеся в ножках мозга.
Функции центров среднего мозга. Передние холмики пластинки четверохолмия выполняют функцию одного из отделов зрительного анализатора. Сюда приходят импульсы от клеток сетчатки глаза. Эта импульсация идет по ответвлениям волокон зрительных нервов, передающих сигнализацию от
ганглиозных клеток сетчатки до ядер таламуса. Таким образом передние бугорки получают как бы копию сигналов, поступающих в таламус. Эта сигнализация обрабатывается нейронами холмиков и от них идет к таламусу. Здесь же замыкается ряд рефлекторных дуг, в частности рефлексы, регулирующие просвет зрачка и аккомодацию глаза. Аккомодацией называют приспособление глаза к видению предметов, находящихся на разном удалении от глаза. Это происходит главным образом путем изменения кривизны хрусталика, выполняющего роль линзы, фокусирующей световые лучи на сетчатке.
Зрачковый рефлекс проявляется сужением диаметра зрачка в ответ на увеличение освещения или открытие глаза. Эфферентные волокна этого рефлекса начинаются от вегетативных нейронов добавочного ядра глазодвигательного нерва и идут к цилиарной мышце и круговым мышцам радужной оболочки глаза. Зрачковый рефлекс легко вызывать и наблюдать. Поэтому он часто используется как для диагностики нарушений функций зрительного анализатора, так и оценки состояния ствола мозга. Исчезновение зрачкового рефлекса свидетельствует о далеко зашедшем угнетении функций ствола мозга. Например, об избыточной глубине наркоза или гипоксии.
Задние холмики пластинки мозга выполняют роль отдела слухового анализатора. Сюда приходят импульсации от ядер слухового нерва. Здесь замыкаются рефлексы, регулирующие сокращение мышц среднего уха, которые защищают структуры внутреннего уха от воздействия избыточной энергии звуковых волн.
В бугорках четверохолмия замыкаются также установочные и ориентировочные рефлексы, возникающие в ответ на действие новых или неожиданных световых и звуковых раздражителей. Эти рефлексы проявляются вздрагиванием, поворотом глаз, головы в сторону раздражителя, настораживанием, изменением позы, подготавливающим организм к так называемому старт-рефлексу или оборонительным реакциям.
Ретикулярная формация среднего мозга принимает участие в интегративных реакциях, обеспечивающих формирование Циклов сна и бодрствования.
Функции ядер III и IV пары черепных нервов тесно связаны с функциями бугорков четверохолмия. Эти ядра участвуют в замыкании рефлексов, регулирующих тонус глазных мышц и Движения глаз.
Ядро глазодвигательного нерва (III пара) иннервирует прямые и нижнюю косую мышцы глаза, сокращение которых вызывает поворот глаз вверх, вниз, к носу и вниз к углу носа. Ин- нервируется также мышца, поднимающая веко. Опускание века регулирует лицевой нерв.
Блоковый нерв (IV пара) иннервирует верхнюю косую мышцу, обеспечивающую поворот глаза вверх-наружу.
Красное ядро. Красное ядро участвует в регуляции тонуса скелетных мышц и движений, обеспечивающих сохранение нормального положения тела в пространстве и принятие позы, создающей готовность к ответной реакции на действие раздражителей.
От красного ядра к спинальным двигательным нейронам идет рубро-спинальный тракт, перекрещивающийся уже на уровне среднего мозга и идущий в боковых канатиках спинного мозга. Волокна этого тракта (посредством вставочных нейронов серого вещества спинного мозга) оказывают возбуждающее влияние на а- и у-мотонейроны сгибателей и тормозят большинство мотонейронов разгибателей.
Роль красного ядра хорошо видна в эксперименте на животных. При перерезке ствола головного мозга (децеребра- ции) на уровне среднего мозга ниже красного ядра развивается состояние, называемое децеребрационной ригидностью. Конечности животного выпрямлены и напряжены, голова и хвост запрокинуты к спине. Это свидетельствует о нарушении баланса между тонусом мышц-антагонистов в сторону резкого преобладания тонуса мышц-разгибателей. В этой ситуации тормозное действие красного ядра и коры мозга на мышцы- разгибатели устранено, а возбуждающее действие на них ядер моста и продолговатого мозга (ядра Дейтерса) сохраняется.
Если перерезка головного мозга проводится между средним и промежуточным мозгом (мезенцифальное животное), то выраженная децеребрационная ригидность не развивается.
У мезенцефального животного проявляются выпрямительные рефлексы. Если положить такое животное на спину, то оно займет обычное положение (встанет на лапы), выпрямится. Выпрямление происходит в два этапа: 1) поворот головы, установление ее теменем кверху (рефлекс запускается с рецепторов вестибулярного аппарата); 2) возврат туловища в ортостатическое, обычное относительно головы, положение (рефлекс запускается с рецепторов мышц шеи и кожной поверхности тела). В осуществлении этих рефлексов участвуют все двигательные центры ствола мозга. Красное ядро важно для установления необходимого баланса между тонусом мышц-сгибателей и мышц-разгибателей.
Позотонические и выпрямительные рефлексы часто называют статическими рефлексами, потому что они обеспечивают нормальную позу и равновесие в покое (в условиях стояния, сидения или лежания).
Выделяют также статокинетическиерефлексы, обеспечивающие равновесие и оптимальную позу при движении (ходьбе, прыжках, вращении, ускорении).
В медицинской практике с диагностической целью особенно часто используются статокинетические рефлексы, возникающие при раздражении рецепторов вестибулярного аппарата вращением. Если человека, находящегося в вертикальном положении, вращать против часовой стрелки, то его глаза будут медленно смещаться вправо (это способствует удержанию взора на одном предмете и восприятию его изображения), затем скачком переместятся в обычное для головы положение.
Такие скачкообразные движения глаз называют глазным нистагмом. На практике проще наблюдать нистагм после остановки вращения (для этого применяют специальное кресло). Быстрый компонент поствращательнного нистагма направлен в сторону, противоположную вращению. Определяя время от момента остановки вращения до прекращения нистагма, судят о возбудимости вестибулярных и мозговых структур, устойчивости человека к вращению и укачиванию. В частности, это важно при профотборе (летчики, моряки, космонавты).
Двигательные центры ствола мозга координируют последовательность выпрямительных и позных движений, обеспечивают условия для выполнения целенаправленных (произвольных движений). Функционирование стволовых центров координируется корой мозга. Это влияние осуществляется как непосредственно по пучку волокон, идущих от моторной зоны коры к красному ядру, так и опосредованно через мозжечок, который посылает пучки эфферентных волокон как к красному, так и к другим ядрам ствола.
Черная субстанция. Нейроны черной субстанции оказывают влияние на нижележащие двигательные и некоторые вегетативные ядра ствола мозга, а также на базальные ганглии (рис. 7.9). Черная субстанция участвует в регуляции тонуса
мышц, позы и движений. Она координирует акты жевания и глотания, влияет на формирование дыхательных движений и кровяное давление.
Между черной субстанцией и базальными ганглиями существуют двусторонние связи. Имеется пучок волокон, проводящий импульсы от полосатого тела к черной субстанции, и до- фаминергический путь, проводящий импульсы в обратном направлении (рис. 7.9). Следовательно, на базальные ганглии черная субстанция воздействует через нейроны, продуцирующие дофамин и использующие его в качестве медиатора. Дофаминергические нейроны оказывают преимущественно тормозное влияние на структуры базальных ганглиев. У некоторых людей при ряде заболеваний, а также в связи с возрастом выработка дофамина в нейронах черной субстанции нарушается. Это приводит к появлению симптомов болезни Паркинсона (см. ниже).
Двигательная
область
Полосато*
тело
Черная
субстанция
Рис.
7.9.
Важнейшие афферентные и эфферентные
связи базальных
ганглиев:
/
— паравентрикулярное ядро;
2
— вентролатеральное ядро; 3 — срединные
ядра, СЯ
—
субталамическое ядро;
4
— кортикоспинальный тракт; 5 —
кортикомосто- вой тракт;
6 —
эфферентный путь от бледного шара к
среднему мозгу
Кора
большого
М0*Г*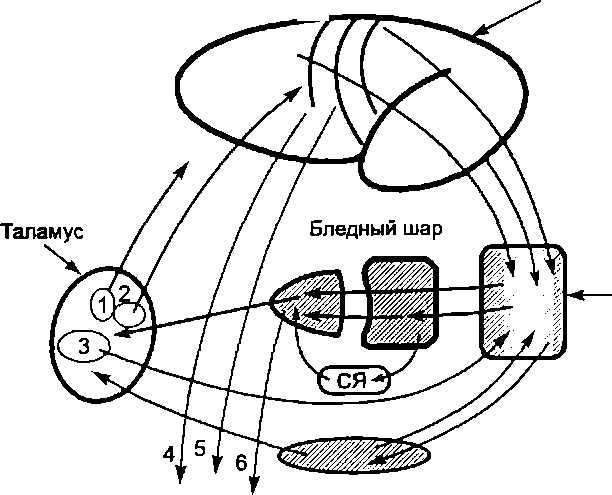
пульсов достигают коры. Таким образом, черная субстанция участвует в замыкании одного из кругов, по которым циркулирует импульсация между корой и подкорковыми образованиями.
7.6. Мозжечок
Основная функция мозжечка — координация произвольных движений, а также тонуса скелетных мышц. Мозжечок может оказывать влияние и на вегетативные процессы.
Мозжечок обеспечивает плавность, соразмерность движений, равновесие тела, участвует в формировании двигательных программ и контроле быстрых комплексных заученных движений, которые после достаточной тренировки могут совершаться автоматически (комплексные упражнения в спорте, игра на музыкальных инструментах). Мозжечок способствует экономии затрат энергии на движение, предотвращая вовлечение избыточного количества мышц. Он облегчает переходы к активации мышц-антагонистов, способствуя тем самым плавности и точности начала и конца движения. Свои влияния на скелетные мышцы мозжечок оказывает через двигательные центры ствола и спинного мозга, а также благодаря двусторонним связям с корой головного мозга. В мозжечок поступает информация от рецепторов мышц, суставов, сухожилий, кожи, вестибулярной, слуховой, и зрительной систем.
Мозжечок также получает информацию о том, какие команды передаются по пирамидным и экстрапирамидным трактам к спинальным двигательным центрам. Эта информация доставляется в мозжечок по ответвлениям волокон пирамидного тракта и эфферентным путям, идущим к мозжечку от моста.
Поступающая в мозжечок информация, перерабатывается на нейронах червя и полушарий мозжечка. Ключевую роль в интегративной функции мозжечка выполняют крупные грушевидные нейроны клетки Пуркинье. Их насчитывают до 30 млн. В этих клетках формируются эфферентные импульса- Ции, которые после переключения на подкорковых ядрах мозжечка направляются к красному, вестибулярным и ретикулярным ядрам ствола мозга. Эта импульсация оказывает преимущественно тормозное избирательное влияние на нейроны Двигательных ядер. Значимость мозжечка для регуляции движений и тонуса мышц становится очевидной при наблюдениях за последствиями повреждений мозжечка.
После повреждения мозжечка выявляются симптомы:
астении быстрая утомляемость и снижение силы мышц;
атаксии — нарушение координации движения. У таких больных неуверенная, шаткая походка с широко расставленными ногами и избыточными движениями-"бросками" из стороны в сторону (симптом, аналогичный таковому при алкогольном опьянении);
астазии, дисметрии и тремора. Астазия и тремор выражаются в потере способности к слитным длительным (тета- ническим) сокращениям. Мозжечковый тремор отмечается не в состоянии покоя, а во время движения. Это одна из причин дисметрии, проявляющейся при выполнении движений. При целенаправленных действиях амплитуда периодических движений становится столь размашистой, что мешает достигать цели движения. Одним из тестов для выявления этих мозжечковых симптомов служит пальце-носовая проба: человек с закрытыми глазами должен прикоснуться пальцем вытянутой руки к кончику носа. При повреждении мозжечка троектория движения руки будет зигзагообразной и не попадающей в цель;
адиадохокинеза замедление скорости смены одной двигательной реакции на другую. Например, человек не способен быстро вращать кистями рук, меняя положения супинация—пронация;
гипотонии — снижение тонуса мышц, особенно ярко выраженное сразу и в начале восстановительного периода после повреждения мозжечка. Если поврежден только червь мозжечка, то часто выявляется дистония — одни мышцы находятся в состоянии гипертонуса, другие — атоничны;
головокружения, при котором выявляется глазной нистагм;
дефектов речи — больной говорит медленно, произнося излишне раздельно не только слова, но и слоги (скандированная речь).
После повреждения мозжечка описанные симптомы нарушений тонуса мышц и движений постепенно исчезают. В этом проявляется пластичность нервных центров, которые могут восстанавливать регуляции, нарушенные из-за повреждения некоторых отделов мозга.
7.7. Ретикулярная формация ствола мозга
Ретикулярная формация образована совокупностью многочисленных нейронов, лежащих отдельно или сгруппированных в ядра (см. рис. 7.7, 7.8). Общим признаком этих нейронов является то, что их отростки сильно ветвятся и образуют многочисленные синаптические контакты как между собой, так и с нейронами других ядер мозга. У нейронов, входящих в ретикулярные ядра, имеются длинные аксоны, формирующие проводящие пути к спинному мозгу, ядрам ствола мозга, мозжечка и таламуса. У большинства остальных ретикулярных нейронов — длинные дендриты и короткий сильно ветвящийся аксон. Эти разветвления образуют своеобразную сеть (ретикулум). Отсюда утвердилось название — ретикулярная система.
Ретикулярные структуры начинают выявляться с верхних сегментов спинного мозга, продолжаются в центральной части шейных сегментов, переходят в ретикулярную формацию ствола мозга и заканчиваются в промежуточном мозге. Нейроны ретикулярной системы ствола располагаются в толще серого вещества продолговатого мозга, моста и среднего мозга.
Афферентные связи ретикулярной формации чрезвычайно многочисленны. Она получает информацию, идущую практически от всех сенсорных рецепторов. Эта информация передается от проводящих путей всех анализаторов за счет ветвления афферентных волокон в проводящих путях. На одном и том же нейроне конвергируют сигналы от разных рецепторов (тактильных, зрительных, слуховых и др.).
В ретикулярных ядрах проводится своеобразный контроль биологической значимости поступающей сигнализации. В зависимости от принятого решения ретикулярная формация может облегчать или тормозить проведение импульсов через нервные центры. Она может активировать как отдельные зоны, так и всю кору. Благодаря этим свойствам ретикулярная формация способна влиять на уровень и направленность внимания, доставку импульсов к различным отделам ЦНС.
Активирующее влияние ретикулярной формации на структуры коры и подкорки осуществляется по восходящим путям, идущим от гигантоклеточного, латерального и вентрального ретикулярных ядер продолговатого мозга, а также ядер моста и среднего мозга. Эти пути доставляют импульсацию к неспецифическим ядрам таламуса. После переключения в таламиче- ских ядрах импульсы приходят к коре. Кроме того, от перечисленных ретикулярных ядер идет сигнализация к заднему гипоталамусу и полосатому телу.
При раздражении некоторых локальных участков ретикулярных структур продолговатого мозга и моста можно вызвать также торможение активности коры и сон. На ЭЭГ при этом возникнут низкочастотные (1 —4 Гц) волны. На основе описанных фактов считают, что важнейшими функциями восходящих влияний ретикулярной формации являются регуляция цикла сон—бодрствование и уровня сознания. Наряду с восходящими активирующими и тормозящими влияниями на высшие отделы головного мозга ретикулярная формация оказывает активирующие и тормозные воздействия по нисходящим путям на центры спинного мозга.
Ретикулярная формация участвует в регуляции сенсорных, двигательных и вегетативных функций.
Регуляция сенсорных функций осуществляется путем влияния на: I) проведение афферентных импульсов и возбудимость; 2) возбудимость рецепторов. Это осуществляется главным образом путем влияния ретикулярной формации на активность симпатических нейронов, иннервирующих органы чувств. В результате может повышаться острота зрения, слуха, тактильная чувствительность. При действии раздражителей, сигнализирующих об опасности, ретикулярная формация активирует не только проводящие пути, но и сенсорные рецепторы.
Ретикулярные ядра ствола мозга входят в структуру жизненно важных отделов дыхательного центра и центров регуляции кровообращения. Ретикулярные нейроны включены в механизмы регуляции всех вегетативных функций.
Ретикулярная формация принимает участие в регуляции движений. На ее ядрах происходит переключение как восходящих путей, идущих от проприорецепторов и спинного мозга к головному мозгу, так и нисходящих двигательных путей от коры мозга, базальных ядер, мозжечка и красного ядра.
Прямое влияние ретикулярной формации на двигательные центры спинного мозга осуществляется: 1) по медиальному ретикулоспиналь- ному тракту, идущему от каудального и орального ядер моста, активирующему а- и у-мотонейроны мышц-разгибателей и тормозящему мотонейроны мышц-сгибателей туловища и конечностей; 2) по латеральному ретикулоспинальному тракту, идущему от гигантоклеточного яд- pa, активирующему а- иу-мотонейроны мышц-сгибателей конечностей и торможение мышц-разгибателей.
Активирующее влияние ретикулярной формации на моторные центры спинного мозга может осуществляться через так называемую у-пет- лю, по аксонам ретикулярных нейронов импульсы приходят к у-мотоней- ронам (см. рис. 7.4), от них — к интрафузальным мышечным волокнам, которые, сокращаясь, активируют рецепторы мышечных веретен. Им- пульсация от этих рецепторов рефлекторно вызывает сокращение соответствующей мышцы.
В центральной части ретикулярного гигантоклеточного ядра обнаружен участок, раздражение которого тормозит все двигательные рефлексы спинного мозга. Наличие тормозных влияний головного мозга на спинной было открыто И.М. Сеченовым в опытах на лягушке. Он делал перерезку спинного мозга по промежуточному мозгу и помещал на разрез кристаллик соли. При этом двигательные спинальные рефлексы не проявлялись или становились ослабленными. Так было впервые выявлено то, что один нервный центр может тормозить активность другого. Это явление назвали центральным торможением.
Ретикулярная формация среднего мозга также координирует функции нейронов в ядрах глазодвигательных черепных нервов (III, IV, VI пары). Благодаря такой координации возможны содружественные движения глаз в вертикальном и горизонтальном направлении, сведение и разведение оптических осей глаз.
7.8. Промежуточный мозг
Промежуточный мозг расположен между средним мозгом и подкорковыми структурами. Он окружает полость третьего желудочка мозга и состоит из таламической области и гипоталамуса (см. рис. 7.8). В таламическую область входит таламус, эпиталамус (эпифиз) и метаталамус (коленчатые тела, их зачастую рассматривают как ядра таламуса).
Таламус. Таламус представляет собой парное образование, составляющее основную массу промежуточного мозга. Он занимает дорсальную часть промежуточного мозга и отделяется от нижележащего гипоталамуса бороздой. От базальных ядер таламус отделяется внутренней капсулой.
В таламусе выделяют до 40 ядер серого вещества. Морфологи их делят по расположению на четыре группы: переднюю, заднюю, латеральную и медиальную. Физиологи выделяют специфические, ассоциативные и неспецифические ядра.
Специфические ядра таламуса (их еще называют релейными, переключательными) обеспечивают быстрое переключение импульсации, идущей практически от всех сенсорных рецепторов (за исключением обонятельных), к сенсорным областям коры. При этом от каждого ядра импульсация передается в отдельные участки коры, выполняющие функцию центрального конца соответствующего анализатора.
Так, в задние вентральные ядра приходит сигнализация от тактильных, вкусовых рецепторов и проприорецепторов. После обработки на переключающих релейных нейронах, у которых мало дендритов и длинный аксон, импульсы идут в соматосен- сорную область коры (в постцентральную извилину, поля 1, 2, 3). Эта область ответственна за формирование ощущений прикосновения, позы тела и тонуса мышц, восприятие схемы тела.
Латеральные коленчатые тела относят к подкорковым зрительным центрам. Сюда приходят импульсы по зрительным трактам и после переключения направляются к затылочной доле коры (поля 17—19). Импульсы от слуховых рецепторов переключаются в медиальных, коленчатых телах, идут к височной коре в область Гешле (поля 41,42).
Вышеперечисленные специфические ядра называют сенсорными. Среди специфических ядер таламуса имеются и несенсорные ядра. Они обеспечивают переключение импульсов не от чувствительных восходящих путей, а от других областей мозга. В таких ядрах, например, переключаются импульсы, идущие от красного ядра, базальных ганглиев и зубчатого ядра мозжечка к моторной зоне коры.
Ряд передних ядер участвует в передаче сигнализации, поступающей от мамиллярных тел, к лимбической системе. Таким образом, эти ядра участвуют в обеспечении круговой циркуляции импульсов по кольцу: лимбическая кора — гиппокамп — гипоталамус — миндалевидное тело — таламус — лимбическая кора. Такую циркуляцию импульсов называют эмоциональным кругом Пепеца. Этот механизм участвует в формировании эмоций. К специфическим ядрам таламуса подходят также проводящие пути от коры, ствола и ретикулярной формации. По этим путям могут передаваться как возбуждающие, так и тормозные влияния на переключающие нейроны. Благодаря таким связям кора мозга способна регулировать потоки идущей к ней информации, блокировать передачу сигналов одной модальности и облегчать передачу другой.
Ассоциативные ядра таламуса (медиодорсальное, латеральное, ядра подушки и др.) отличаются тем, что к их нейронам приходят импульсы, уже обработанные в других нервных центрах и ядрах таламуса. Характерно, что на один и тот же нейрон приходят импульсы разных модальностей. Например, от центров, обеспечивающих зрительную, тактильную и болевую чувствительность. Нейроны ассоциативных ядер являются полисенсорными и обеспечивают возможность интегратив- ных процессов, в результате которых формируются сигналы, передаваемые в ассоциативные области коры мозга. Эти импульсы обеспечивают проявление таких психических процессов, как узнавание предметов и явлений, согласование речевых, зрительных и двигательных функций, формирование представления о позе и положении тела.
Неспецифические ядра таламуса представлены центральными и парафасцикулярными ядрами, а также ретикулярным ядром таламуса. Эти ядра содержат мелкие нейроны, образующие многочисленные синаптические связи с нейронами как таламуса, так и лимбической системы, базальных ядер, гипоталамуса, ствола мозга. По чувствительным восходящим путям сюда приходит сигнализация от болевых и температурных рецепторов, а по сетям нейронов ретикулярной формации практически от всех рецепторных входов.
Эфферентные пути от неспецифических ядер идут ко всем зонам коры как непосредственно, так и через другие талами- ческие и ретикулярные ядра. Имеются также нисходящие пути к стволу мозга. Неспецифические ядра благодаря своим многочисленным связям обеспечивают взаимодействие и координацию работы отделов головного мозга. Они оказывают модулирующее влияние на состояние нервных центров, обеспечивают оптимальную рабочую настройку этих органов.
Таламус выполняет также проводниковую и релейную функции. В нем переключаются, обрабатываются и проводятся все импульсы, идущие от рецепторных входов, спинного мозга, ствола и мозжечка, к коре мозга. В таламусе заканчивается ряд нисходящих путей от коры, лимбической системы и базальных ганглиев.
Таламус может обеспечить координацию ряда сложных Двигательных актов: сосание, жевание, глотание, смех.
Таламус иногда называют центром болевой чувствитель- ности. Это мнение сложилось на основе наблюдений о том, что разрушение ряда специфических ядер и структур подушки приводит к исчезновению некоторых сильных, стойких болей, которые не поддаются снятию другими средствами. Таламус участвует также в формировании эмоций и психических процессов, обеспечивающих узнавание, обучение, память.
Гипоталамус. Гипоталамус занимает вентральный отдел промежуточного мозга. Он лежит ниже таламуса, образуя стенки нижней части третьего желудочка. В гипоталамусе выделяют серый бугор с воронкой и нейрогипофизом, сосцевидные тела. Нижняя часть гипоталамуса ограничена средним мозгом, передневерхняя — передней спайкой, терминальной пластинкой и зрительным перекрестом. Последний образован волокнами зрительных нервов, медиальная часть которых переходит на противоположную сторону и формирует зрительный тракт.
В составе гипоталамуса выделяют до 50 ядер. Среди наиболее значимых (см. рис. 7.8) — преоптическое, паравентрику- лярное, супраоптическое, переднее, вентро- и дорсомедиаль- ные и задние ядра.
Гипоталамус имеет множественные афферентные и эфферентные связи со структурами ствола, промежуточного, конечного и спинного мозга. Это позволяет гипоталамусу выполнять интеграцию вегетативных соматических и эндокринных регуляций в организме. Взаимодействие с эндокринными железами основывается на особых свойствах нейронов преоптиче- ской и передней областей, а также вентромедиального и инфун- дибулярного ядер гипоталамуса. Область расположения этих ядер называют гипофизотропной зоной. Нейроны этой зоны секретируют так называемые рилизинг-факторы — вещества, которые через систему воротной вены гипофиза передаются аденогипофизу и регулируют секрецию им тропных гормонов, управляющих секреторной функцией эндокринных желез.
На количество выделяемого гипоталамическим нейроном медиатора (нейрокринную активность) оказывают влияние:
нервные импульсы, приходящие от лимбической и ретикулярной системы, миндалины и других ядер гипоталамуса;
уровень гормонов и других биологически активных веществ в крови. Последнее становится возможным благодаря очень высокой проницаемости гематоэнцефалического барьера в области гипоталамуса. В этой области сосредоточено большое количество кровеносных капилляров (на 1 мм3 — Д° 2600, а в затылочной доле коры — только 900), обеспечивающих интенсивный кровоток по сравнению с другими областями мозга.
Благодаря особенностям кровоснабжения и проницаемости сосудов нейроны гипоталамуса могут получать информацию о составе внутренней среды организма. Некоторые из этих нейронов имеют свойства рецепторов. В частности, есть нейроны, чувствительные к уровню осмотического давления, электролитного состава, температуре крови.
Гипоталамус выполняет важнейшую роль в регуляции гомеостаза. Животное с поврежденным гипоталамусом погибает из-за невозможности сохранения гомеостаза.
Свои влияния на вегетативные функции гипоталамус реализует не только через гипоталамо-гипофизарно-эндокрин- ную систему, но и путем регуляции тонуса симпатических и парасимпатических центров ствола и спинного мозга.
Гипоталамус считают высшим центром вегетативных регуляций. При раздражении переднего гипоталамуса активируются преимущественно парасимпатические центры, при раздражении заднего — симпатические. Но эти реакции не всегда однозначны. Исследователи приходят к выводу о существовании тесной связи и взаимообусловленности функций переднего, среднего и заднего гипоталамуса, который в ряде случаев выступает как единый интегративный центр.
В гипоталамусе располагаются центры регуляции теплопродукции, теплоотдачи и поддержания температурного гомеостаза. Обнаружены центры регуляции пищевого поведения (голода и насыщения), биологических ритмов, сна и бодрствования, эмоциональных реакций: ярости, гнева, страха.
Гипоталамус участвует в регуляции энергетического обеспечения движений путем влияния на вегетативные функции (кровоток, дыхание и другие) и приспособления их к потребностям мышечной системы.
7.9. Конечный мозг
Конечный мозг (telencephalon) состоит из двух полушарий большого мозга (больших полушарий).
В конечном мозге по морфологическим и функциональным Признакам выделяют кору больших полушарий (КБП), лимби- Ческую систему и базальные ганглии.
Базальные ганглии. Базальные ганглии, или подкорковые ядра, располагаются в глубине больших полушарий между их лобными долями и промежуточным мозгом. Это парные образования, состоящие из ядер серого вещества, разделенных прослойками (внутренней и наружной капсулами) белого. В структуры базальных ганглиев входят: полосатое тело, состоящее из хвостового ядра и скорлупы, бледный шар и ограда (см. рис. 7.9). Они представляют собой функционально единое образование.
По функциональным показателям к системе базальных ганглиев относят также черную субстанцию и субталамиче- ское ядро, а в некоторых случаях и миндалевидное тело.
Ряд сведений о роли базальных ганглиев удалось получить, изучая ответные реакции на раздражение этих структур электрическим током. При электрическом раздражении полосатого тела возникают относительно простые движения: сгибание конечностей противоположной стороны тела, поворот головы в сторону. Отмечается также появление признаков сна (торможение ориентировочных реакций, возникновение медленных волн на электроэнцефалограмме), может исчезнуть ощущение боли. Электрическое раздражение бледного шара и ограды вызывает сокращение мышц шеи, лица, конечностей, жевательные и глотательные движения. Обнаружено также влияние этих структур на функции внутренних органов, проявление пищедобывательных реакций, образование и сохранение условных рефлексов.
При ревматическом поражении базальных ганглиев наблюдаются гиперкинезы — непроизвольные стереотипные повторяющиеся движения с разным ритмом и амплитудой (хорея, атетоз). Эти движения особенно выражены у конечностей и проявляются в покое. Нарушается также регуляция тонуса скелетных мышц.
Важное значение в понимании роли функционирования базальных ганглиев имело выяснение их афферентных и эфферентных связей.
Афферентные волокна идут главным образом к полосатому телу и передают импульсы от трех источников: всех отделов коры мозга, таламуса и черной субстанции (см. рис. 7.9). После обработки в полосатом теле эти импульсы идут к бледному шару и черной субстанции. Между нею и полосатым телом имеются двусторонние связи.
Черная субстанция влияет на нейроны базальных ганглиев с помощью медиатора дофамина. При дегенерации дофами- нергических нейронов, нарушении синтеза и выделения этого медиатора нарушаются и функции системы базальных ганглиев. Появляются симптомы заболевания, называемого паркинсонизмом (дрожательный паралич). Среди симптомов паркинсонизма выделяют гипокинезию — резкое затруднение двигательных реакций, особенно их начала. Для таких больных характерна осторожная (шаркающая) походка мелкими шажками. Появляется тремор покоя (частота 4—7 Гц) особенно выраженный на кистях рук. Этот тремор исчезает при осуществлении произвольного движения (в отличие от тремора, вызванного повреждением мозжечка, проявляющегося во время движений).
Выявление роли дофамина в этом заболевании позволило найти относительно эффективный способ его лечения путем систематического введения таким больным вещества L-Дофа, из которого образуется дофамин. Сам дофамин в этом плане оказался неэффективным, так как гематоэнцефалический барьер для него непроницаем.
Эфферентные импульсы от полосатого тела идут к бледному шару и черной субстанции, а от них — к таламусу и далее к двигательной области коры мозга. Бледный шар передает также импульсации к двигательным ядрам ствола мозга и субталамическому ядру.
Благодаря многосторонним связям базальных ганглиев создается несколько кругов циркуляции импульсов между корой и подкорковыми структурами, которые обеспечивают регуляцию целенаправленного поведенческого акта. При этом в коре мозга (прежде всего в ассоциативной) создается замысел движения, а базальные ганглии участвуют в формировании программы движения. Эта программа через таламус передается в моторную зону коры, а оттуда по пирамидным путям — к спи- нальным мотонейронам и мышцам.
Таким образом, базальные ганглии участвуют в регуляции сложных целенаправленных движений. Они обеспечивают формирование программ движения, совместно с мозжечком производят коррекцию амплитуды, силы, скорости сокращения мышц, а также их тонуса. Особенно необходимо участие базальных ганглиев в формировании стереотипных медленных червеобразных движений. Базальные ганглии принимают участие также в регуляции цикла сон—бодрствование, реализации некоторых инстинктов и условных рефлексов.
Лимбическая система и ее функции. Лимбическая система состоит из филогенетически старых отделов переднего мозга. В названии (limbus — край) отражена особенность ее расположения в виде кольца между новой корой и конечной частью ствола мозга. К лимбической системе относят медиально расположенные структуры конечного мозга: поясную, пара- гиппокампальную и зубчатую извилины, гиппокамп, обонятельную луковицу, обонятельный тракт и прилежащие к нему участки коры. Кроме того, после выяснения функциональных связей перечисленных структур с другими отделами головного мозга к лимбической системе стали относить также миндалину, переднее и септальные таламические ядра, гипоталамус и мамиллярные тела (рис. 7.10).
Лимбическая система имеет множественные афферентные и эфферентные пути.
От височной области к лимбической системе доставляется информация, проанализированная и переработанная в зрительной, слуховой и соматосенсорной зонах коры. Эта сигнализация проводится к миндалине и гиппокампу. Она имеет важное значение для возникновения и запуска эмоциональных
и
ствола мозга:
А
— круг Пепеца;
Б —
круг через миндалину;
МТ — мамиллярные тела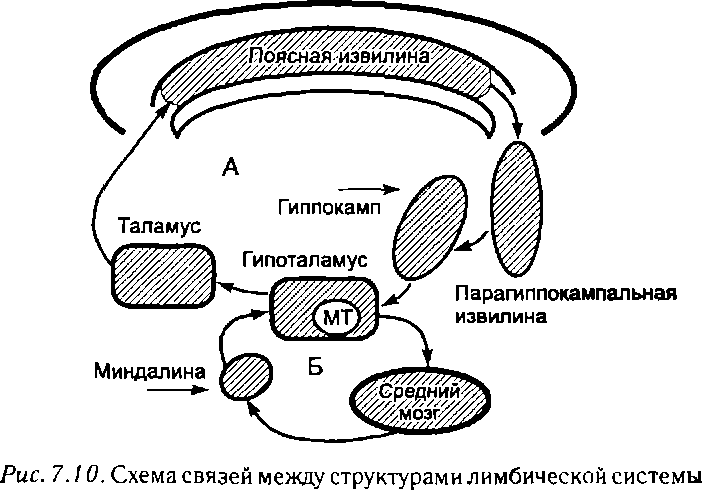
реакций, которые формируются на основе функционирования лимбической системы.
Лобная область коры оказывает корригирующее действие на функции лимбической системы. Она сдерживает проявление излишних эмоциональных реакций, связанных с удовлетворением простейших биологических потребностей и, видимо, способствует появлению эмоций, связанных с реализацией социальных взаимоотношении и творчества.
Эфферентные влияния лимбической системы на соматические и вегетативные функции осуществляются через ма- миллярные тела и другие центры гипоталамуса.
Влияние лимбической системы на деятельность коры (высшие психические функции) осуществляется через эфферентные пути, идущие преимущественно к ассоциативной коре.
В формировании эмоций важная роль принадлежит наличию замкнутых кругов циркуляции нервных импульсов между образованиями лимбической системы. Особую роль в этом играет так называемый лимбический круг Пепеца (гиппокамп — свод гипоталамус — мамиллярные тела — таламус — поясная извилина — парагиппокампальная извилина — гиппокамп). Циркулирующие здесь импульсации иногда называют потоком эмоций.
Другой круг (миндалина — гипоталамус — средний мозг — миндалина) важен в регуляции агрессивно-оборонительных, сексуальных и пищевых поведенческих реакций и эмоций.
Лимбическая система обеспечивает согласование процессов, регулирующих вегетативные, соматические и психические реакции. Важнейшей функцией этой системы является сохранение гомеостаза, обеспечение поведенческих реакций, направленных на сохранение жизни индивидуума и вида. Лимбическая система обеспечивает формирование эмоций и мотиваций.
Лимбические структуры важны для обучения, памяти, выработки и сохранения условных рефлексов. Они участвуют в регуляции внимания, восприятия, запоминания и воспроизведения информации. Особую роль в механизмах памяти играет гиппокамп. Его нейроны обладают сильно выраженным свойством на длительное время увеличивать возбудимость и способность к проведению возбуждения в синапсах вслед за предыдущим раздражением (это называется посттетанической Потенциацией). Это свойство в сочетании с наличием замкнутых кругов циркуляции возбуждения важно для формирования Долговременной памяти.
7.10. Кора полушарий большого мозга
Кора полушарий большого мозга (кора больших полушарий — КБП, или просто кора) представляет собой слой серого вещества, покрывающего большие полушария (рис. 7.11, 7.12). Нейроны в нем располагаются горизонтальными слоями. Толщина этого слоя в разных участках коры колеблется от 1,3 до 4,5 мм. Она максимальна в передней центральной извилине.
В участках коры, относящихся к лимбической системе, функции которой рассмотрены выше, имеются зоны с трехслойным и пятислойным расположением нейронов в структуре серого вещества. Эти участки (филогенетически древней коры) занимают около 10% поверхности полушарий мозга, остальные 90% составляют новую кору.
Структурно-функциональная характеристика новой коры большого мозга. Кора головного мозга регулирует целенаправленные поведенческие реакции человека, его пространственно- временные взаимоотношения с окружающей средой. Она обеспечивает мышление, сознание и познавательные возможности человека, является филогенетически новым, молодым образованием, которое в значительной мере подчинило себе другие структуры мозга, оказывая координирующие влияния на их функции. Такое влияние коры мозга на регуляторные процессы в организме называют кортикализацией функций.
9
17
Рис.
7.1I.
Схема полей коры большого мозга по
Бродману. Цифрами указаны номера полей
и область их локализации. Границы
некоторых полей выделены оттенками
окраски
Центральная
борозда
Премоторная
зона
Лобная
доля
Моторный
центр речи Брока
Центр
слуха (высота тона)
Височная
доля
Мозжечок
Рис.
7.12.
Кора полушарий большого мозга и
некоторые из ее зон и центров
Соматосенсорная
зона
Теменная
доля
Зона
навыков письма
Восприятие
письменной речи
Затылочная
доля
Соматомоторная
зона
Центральное
зрительное поле
Центр
Вернике (восприятие устной речи)
Новая кора имеет шестислойное расположение нейронов. Нейроны разных слоев различаются по цитологическим признакам и функциональным свойствам.
Молекулярный слой. Является самым поверхностным. В нем мало нейронов, но очень много ветвящихся дендритов — отростков пирамидных нейронов, лежащих в более глубоких слоях.
Наружный зернистый слой. Его составляют многочисленные мелкие нейроны самой разной формы. За счет отростков клеток этого слоя обеспечиваются кортико-кортикалъные связи.
Наружный пирамидный слой. Состоит из пирамидных нейронов средней величины, которые наряду с нейронами второго слоя обеспечивают кортико-кортикальные связи между соседними областями коры.
Внутренний зернистый слой. По виду клеток и расположению волокон подобен второму слою. Здесь проходят пучки проводящих воло
кон, связывающих различные участки коры. К нейронам этого слоя приходят импульсации от специфических ядер таламуса.
Внутренний пирамидный слой. Образован средними и крупными пирамидными нейронами. В двигательной области коры эти нейроны особенно крупные (так называемые гигантские, диаметром 50— 100 мкм, пирамидные клетки Беца). Аксоны этих клеток составляют быстропрово- дяшие (до 120 м/с) волокна пирамидного тракта.
Слой полиморфных клеток. В нем чаще всего встречаются клетки, аксоны которых образуют кортико-таламические пути.
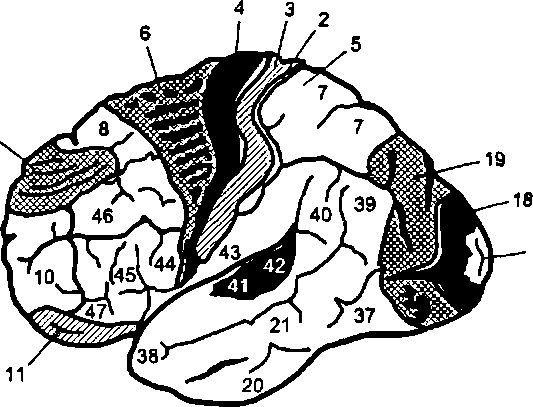
Нейроны 1 —4-го слоев участвуют преимущественно в восприятии и переработке приходящих к коре импульсов и перераспределении сигналов между корковыми центрами. На нейронах 5—6-го слоев формируются эфферентные нисходящие импульсации. В нейронном составе и цитологических особенностях разных участков коры имеются значительные отличия. По этим отличиям Бродман разделил кору на так называемые поля (см. рис. 7.10). В настоящее время выделяют до 53 корковых полей. Оказалось, что многие из полей, выделенных на основе гистологических данных, совпадают по топографии с физиологическими функциональными центрами.
Белое вещество полушарий головного мозга образовано нервными волокнами. Выделяют ассоциативные волокна, передающие импульсы к нейронам рядом лежащих извилин (дугообразные волокна), и длинные волокна, доставляющие импульсацию к более удаленным участкам одноименного полушария. Такие волокна объединяются в продольные пучки.
Комиссуральные волокна — поперечные волокна, передающие импульсы между левым и правым полушариями.
Проекционные волокна — проводят импульсы между корой и другими отделами мозга.
Перечисленные виды нервных волокон участвуют в создании нейронных цепей с нейронами, расположенными на значительных расстояниях. В коре обнаружен также особый видлокальных нейронных цепей, образованных рядом лежащими нейронами. Эти структуры получили название функциональных кортикальных колонок. Нейронные колонки образованы группами нейронов, расположенных друг над другом перпендикулярно к поверхности коры. Диаметр функциональной колонки — до 1 мм. Нейроны одной функциональной колонки получают импульсы от одного и того же афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи, по которым обмениваются информацией. Таким образом увеличиваются надежность регуляции и возможности анализа информации, приходящей в корковые проекционные зоны.
Особенностью корковых функциональных полей является также их соматотопическая организация. Участки кавдого рецептивного поля представлены в проекционных областях коры не хаотично. Они в определенной мере копируют последовательность расположения рецепторов на периферии.
Наблюдается также экранная организация ряда корковых центров. Внешнее рецепторное поле проецируется как бы на развернутую поверхность центра. Однако эта поверхность не однослойная, а чаще всего сформирована группами лежащих друг за другом нейронов, объединенных в колонки — своеобразные функциональные ячейки экранного центра.
В ряде сенсорных и моторных зон коры выделяют центральное проекционное поле, обеспечивающее восприятие ощущений определенных модальностей (свет, звук, прикосновения, температура), и вторичные проекционные поля. Функцией последних является обеспечение понимания связи первичного ощущения с другими предметами и явлениями окружающего мира.
Зоны представительства рецептивных полей в коре в значительной мере перекрываются. Отмечается пластичность нервных центров коры, возможность перестройки специализации и восстановление функций после повреждения какого- либо из центров за счет соседних корковых структур. Особенно выражены компенсаторные возможности нервных центров в детском возрасте.
Считается, что кора мозга, формируясь на поздних этапах эволюционного развития высших организмов, в определенной мере подчинила себе все нижележащие отделы ЦНС и может оказывать корригирующие влияния на функции этих отделов.
Функциональные области коры полушарий большого мозга. По функциональному признаку в коре выделяют сенсорные, ассоциативные и двигательные области.
Сенсорные области коры. Сенсорные (чувствительные, проекционные) области коры состоят из зон, содержащих нейроны, активация которых импульсами от сенсорных рецепторов или прямым воздействием раздражителей вызывает появление четких ощущений. Эти зоны имеются в затылочной (поля 17—19), теменной (поля 1—3) и височной (поля 21—22, 41—42) долях коры.
В затылочной доле за шпорной бороздой находится первичная зрительная зона (поле 17, занимающее участки коры клиновидной извилины и язычковой дольки). Сюда приходят импульсы от рецепторов сетчатки. Сетчатка глаза представлена в этой зоне соматотопически. Следовательно нейроны, к которым приходит сигнализация от фоторецепторов, расположены по отношению друг к другу подобно рецепторам в сетчатке глаза (правда, эта соматотопичность не абсолютная, область желтого пятна сетчатки имеет относительно большую зону представительства в коре).
Раздражение нейронов коры поля 17 приводит к возникновению световых ощущений. Благодаря этому полю осуществляется восприятие изображения предметов. Следствием его разрушения является потеря зрения.
К. полю 17 примыкает кора вторичной зрительной области, занимающая поля 18—19. Нейроны этих полей полисенсор- ные. К ним приходит импульсация не только от зрительных, но и от тактильных, слуховых и других рецепторов.
Информация о сигналах, поступивших в сенсорные зоны коры, передается также в ассоциативную область коры.
В левую и правую зрительные зоны каждого полушария проецируются одноименные половины сетчатки, в зрительную зону левого полушария приходит импульсация-от левых частей сетчатки каждого глаза (от латеральной половины левого глаза и назальной половины правого глаза).
Слуховая сенсорная кора (поля 41 — 42) расположена в латеральной борозде в области извилияы Гешля. Благодаря им- пульсациям от слуховых рецепторов здесь формируются звуковые ощущения. Определяются характеристики высоты, тембра, громкости звука. В верхней и средней височных извилинах (поля 21—22) находится также зона вестибулярного анализатора. Поступающая сюда сигнализация от рецепторов вестибулярного аппарата обеспечивает формирование ощущений положения тела в пространстве и ускорений. Эта область взаимодействует с мозжечком (через височно-мосто-мозжечковый путь) и участвует в регуляции равновесия тела, коррекции позы и движений. На основе взаимодействия этой области с соматосенсорной и ассоциативной корой происходит осознание схемы тела.
Обонятельная область занимает кору крючка (поля 28, 34), относящуюся к лимбической системе и имеющую трехслойное расположение нейронов. Ее повреждение ведет к аносмии — потере обоняния. При искусственном раздражении этой области возникают ощущения различных запахов по типу галлюцинаций.
Проекция вкусовой чувствительности приходится на поле 43 гиппокампальной извилины.
Соматосенсорная область коры занимает постцентральную извилину (см. рис.7.11, 7.12) теменной доли (поля 1—3), включая парацентральную дольку на медиальной стороне полушарий. В соматосенсорную область приходят импульсы от рецепторов кожи (тактильная, температурная, болевая чувствительность), проприорецепторов (мышечных, суставных сумок, сухожилий) и интерорецепторов (внутренних органов). Из-за перекреста афферентных путей в соматосенсорную зону левого полушария приходит сигнализация от правой стороны тела, а в правое полушарие — от левой. В этой сенсорной зоне также имеется соматотопическое представительство областей тела. Правда, проекции наиболее важных рецептивных зон (пальцев рук, губ, кожи лица, языка, гортани) занимают относительно большие площади, чем проекции таких поверхностей тела, как спина, передняя часть туловища, ноги.
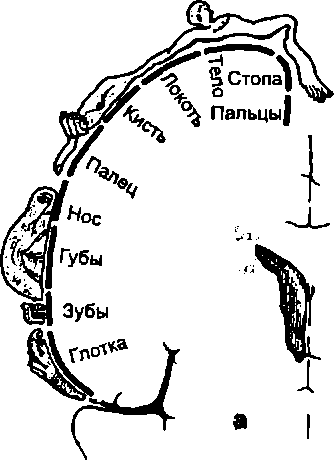
Рис.
7.13.
Представительство чувствительности
частей тела в постцентральной
извйййне
(а)
и двигательных
эойв
прецентральной извилине(б)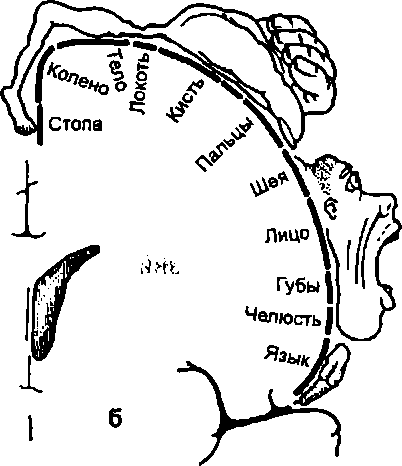
этом чувствительность голеней и стоп проецируется на кору парацентральной дольки медиальной поверхности полушарий.
Знание топографии представительства чувствительности используется в клинике для определения области и степени повреждения коры. При односторонних повреждениях (инсульты, травмы)участка постцентральной извилины выпадают некоторые виды чувствительности соответствующих участков кожи противоположной стороны тела.
Кору постцентральной извилины относят к соматосенсорной области I, выделяют также соматосенсорную область II. Функции и связиэтой зоны менее изучены, чем области I. Она располагается в конце постцентральной извилины на верхней стенке сильвиевой борозды, в верхней теменной дольке (поле 7).
Полагают, что соматосенсорная область II обеспечивает интеграцию ощущений разных сторон тела и координацию движений при одновременном участии мышц обеих сторон тела (в частности, ощупывание обеими руками, хватание, однонаправленное движение обеими руками). Функционирование этой области необходимо для узнавания предметов на ощупь и определения пространственного расположения этих предметов.
Двигательные области коры. Около 130 лет тому назад при воздействии точечными раздражениями электрическим током на кору мозга было обнаружено, что действие на поверхность передней центральной извилины вызывает сокращения мышц противоположной стороны тела. Так была открыта одна из моторных зон коры мозга.
Первичная моторная область коры (поле 4) занимает переднюю центральную извилину (см. рис. 7.12). Имеется соматотопическая организация представительства разных мышечных групп в коре. При этом представительство мышц ног и туловища занимает верхние (прилежащие к срединной борозде) участки извилины и относительно малую площадь, а мышц рук, особенно пальцев, лица, языка и глотки — нижние участки и большую площадь (см. рис. 7.13). В двигательной коре относительно большую площадь занимают те нейронные группы, которые управляют мышцами, осуществляющими разнообразные точные, мелкие, тонко регулируемые движения.
Вторичная моторная область (М2) прилегает к первичной моторной области и располагается на боковой поверхности межполушарной щели.
При повреждении моторных зон М1 и М2 коры утрачивается способность к тонким координированным движениям. Точечные раздражения дополнительной моторной зоны сопровождаются активацией не отдельных мышц, а целой группы мышц, вызывающих направленное движение в суставах. Эти наблюдения позволили сделать вывод о том, что в коре представлены не столько мышцы, сколько движения.
Важная роль в регуляции движений принадлежит также обширным участкам лобной доли коры мозга, расположенной перед центральной извилиной. Это премоторная кора (поле 6) и дополнительная моторная область (поле 8), располагающаяся в верхней лобной извилине.
Премоторная область коры выполняет высшие двигательные функции, связанные с планированием и координацией произвольных движений. Одной из самых сложных проблем в физиологии является выяснение механизма возникновения побуждения к действию, формирования замысла движения и его реализации.
В настоящее время о конкретных механизмах, реализующих превращение замысла движения в программу двигательной реакции (залпы импульсов, посылаемых к мышцам), известно немного. Считается, что замысел движения формируется благодаря функциям ассоциативной и премо- торной коры, взаимодействующих со всеми структурами головного мозга.
В этой области располагаются моторные центры, обеспечивающие движения, связанные со второй сигнальной системой человека (см. рис.7.11, 7.12). Центр письменной речи находится в задней части средней лобной извилины (поле 6), моторный центр устной речи (Брока) — в нижней лобной извилине (поле 44). При повреждении этих центров человек теряет способность соответственно к пониманию и написанию слов, а также к произнесению слов и связной речи.
Информация о замысле движения передается в двигательные области лобной коры. Двигательная кора через нисходящие пути активирует две системы, в которых возможна выработка новых двигательных программ или использование старых, уже отработанных на практике и хранящихся в памяти. Это системы базальных ганглиев и мозжечка. Они в определенной мере дублируют друг друга, хотя и имеют ряд отличий в своих функциях. После проработки в мозжечке и базальных ганглиях импульсы, в которых закодированы программы движения, передаются через ядра таламуса в моторные зоны, прежде всего в первичную моторную область коры. Считается, что эта моторная область выполняет функции не планирования, а выполнения движений. Именно через нее программа Движения, в которой закодирован выбор мышц, последовательность смены их сокращения и расслабления, передается на спинальные мотонейроны и мотонейроны черепных нервов — эти конечные командные пункты, непосредственно вызывающие сокращения мышц.
Влияние моторной коры на сокращения и тонус мышц осуществляется через пирамидные и экстрапирамидные пути. Пирамидные пути начинаются от пирамидных нейронов (гигантских пирамидных клеток Беца, лежащих в 5-м слое, и мелких пирамидных клеток других слоев).
В состав пирамидных кортикоспинальных путей входят аксоны не только нейронов первичной моторной области (поле 4), но и премоторной (поле 6) и соматосенсорной (поля 1, 2, 3).
Абсолютное большинство волокон пирамидного тракта, идущих к спинному мозгу, перекрещивается. В области пирамцд продолговатого мозга — 70%, в шейных и грудных сегментах спинного мозга — остальные 30%. Таким образом, каждая сторона полушарий большого мозга контролирует мышцы противоположной половины тела. Исключение из этого правила составляют мышцы головы, иннервируемые нейронами двигательных ядер черепных нервов (III — VII, IX — XII пары). Лишь часть волокон, идущих от коры к этим ядрам, переходит на другую сторону. В результате каждая сторона двигательной коры может вызывать сокращение мышц обеих сторон головы.
Волокна пирамидного тракта могут оканчиваться синапти- ческими структурами прямо на а- и у-мотонейронах спинного мозга, но большинство этих волокон передает влияние на спинальные мотонейроны, через один или несколько вставочных нейронов. Пирамидный тракт оказывает возбуждающее влияние преимущественно на мотонейроны мышц-сгибателей. На мышцы-разгибатели моторные зоны коры влияют через экстрапирамидные пути.
К экстрапирамидной системе относят двигательные ядра ствола мозга и отходящие от них двигательные пути. В эту систему функционально включаются также мозжечок и базальные ганглии.
Ассоциативные области коры. У человека ассоциативная кора занимает около 50% всей коры большого мозга. Она располагается в участках между сенсорными и двигательными областями. Ассоциативная кора не имеет четких границ со вторичными сенсорными областями как по морфологическим, так и по функциональным признакам.
Ассоциативная кора играет первостепенную роль в формировании высших психических функций человека. Она обеспечивает формирование целевых установок и программ осознанных поведенческих реакций, узнавание и смысловую оценку предметов и явлений, понимание речи, логическое мышление.
Нейроны ассоциативной коры являются полисенсорными. Один и тот же нейрон может возбуждаться при действии нескольких различных (световых, звуковых, тактильных и др.) раздражителей.
Можно считать общепризнанным наличие некоторой специализации в выполнении функций ассоциативными и сенсорными областями коры левого и правого полушарий. Различают психическую, моторную и сенсорную составляющие функциональной асимметрии больших полушарий. Самым ярким свидетельством наличия такой асимметрии является то, что в левом полушарии большинства людей (у 95% правшей и 70% левшей) находятся сенсорный и моторный центры речи.
Полушария, в которых локализуются речевые и другие односторонне представленные двигательные центры, называют доминантными. У абсолютного большинства правшей доминантным является левое полушарие. Оно обеспечивает прежде всего абстрактно-логическое мышление, а правое — конкретно-образное мышление.
Следует отметить, что нет абсолютной предопределенности того, что у человека доминантным станет левое полушарие. По крайней мере, у 30% левшей центры речи, письма и других сложных двигательных актов располагаются в правом полушарии. Если в раннем детстве повреждзется левая половина полушария в зоне центров речи, то постепенно центры формируются в симметричных областях правого полушария и происходит восстановление речи. Это один из примеров пластичности и динамической локализации функций в коре мозга. У взрослого человека после повреждения центров речи полной компенсации функций за счет структур противоположного полушария не происходит.
193
7 Зак. 181
Моторный центр речи находится в нижней лобной извилине (поля 44—45), его также называют центром Брока.
Больные с поражением центра Брока страдают моторной афазией — нарушением артикуляции и построения речевых фраз. Они понимают обращенную к ним речь, но если пытаются говорить сами, то могут произносить лишь очень привычные короткие названия предметов или переходят на произнесение повторяющихся слогов (та-та-та, ни-ни-ни). Теряется также способность к составлению и воспроизведению музыкальных фраз. Характерно, что центр моторной речи соседствует с отделом прецентральной извилины, контролирующим функции мышц языка, рта, глотки, лица. Нарушения моторной речевой функции могут наступать также при повреждении вторичной моторной области коры М2. Раздражение этой области электрическим током сопровождается временной моторной афазией.
Иной тип афазии (сенсорная) развивается при повреждении задних отделов верхней височной извилины — центра Вер- нике. При повреждении этого сенсорного речевого центра человек перестает понимать смысл слов и фраз, не воспринимает речь. При этом способность к произнесению речевых фраз и моторная речь у него сохраняется.
Зона восприятия письменной речи занимает угловую извилину височной доли (поле 39). При ее повреждении утрачивается способность воспринимать написанный текст.
Зона навыков письма располагается в надкраевой извилине нижней теменной дольки (поле 40) доминирующего полушария. При повреждении этого поля теряется способность производить сложные движения, полученные в результате обучения, и, в частности, утрачиваются навыки письма (аграфия).
Учитывая некоторую специализацию функций каждого полушария, следует иметь в виду, что они дополняют друг друга и обеспечивают единство в осуществлении мыслительных процессов и целенаправленных двигательных реакций.
7.11. Автономная (вегетативная) нервная система
Нервную систему организма человека и животных подразделяют на соматическую (анимальную) и автономную (вегетативную).
Соматическая нервная система обеспечивает иннервацию и регуляцию активности скелетной (соматической, поперечнополосатой) мускулатуры, а также восприятие информационных сигналов (раздражений) из внешней среды.
Автономная нервная система (АНС) обеспечивает иннервацию и регуляцию функций сосудов (кровеносных и лимфатических), желез (экзо- и эндокринных) и других внутренних (висцеральных) органов, а также трофическую иннервацию скелетной мускулатуры, органов чувств и самой центральной нервной системы. Основными функциями АНС являются поддержание постоянства внутренней среды организма (гомеостаза) и вегетативное обеспечение различных форм психической и физической деятельности человека или животного.
Морфофункциональные и нейрохимические особенности АНС. Строение, функциональные свойства АНС, механизмы передачи возбуждения с вегетативных нервных волокон на иннервируемые клетки существенно отличаются от таковых соматической нервной системы.
Анатомические и функциональные особенности. В АНС выделяют центральную и периферическую части. Все образования АНС делят на три уровня. Первый уровень — периферическая часть АНС — вегетативные ганглионарные нейроны. Второй уровень — вегетативные преганглионарные нейроны ствола мозга и спинного мозга. Третий уровень — высшие вегетативные центры ЦНС — нейроны гипоталамуса, ретикулярной формации ствола мозга, мозжечка, миндалины, коры больших полушарий. Нейроны третьего уровня регулируют активность как АНС, так и соматической нервной системы.
Важной особенностью АНС является расположение эфферентных (ганглионарных) нейронов за пределами спинного и головного мозга в вегетативных ганглиях или сплетениях. Аксоны этих ганглионарных нейронов называются постганглио- нарными волокнами. В соматической же нервной системе эфферентные нейроны (мотонейроны) располагаются в передних рогах серого вещества спинного мозга или в двигательных ядрах черепных нервов.
Следующая особенность — наличие многочисленных вегетативных ганглиев и нервных сплетений, состоящих из микроганглиев, в периферической части АНС. Число нейронов в них превышает число клеток в спинном мозге. Вегетативные ганглии содержат четыре типа нейронов: эфферентные, афферентные (сенсорные, чувствительные), вставочные (ассоциативные) и клетки-осцилляторы, обладающие автоматией (способностью к самовозбуждению). Вегетативные ганглии могут выполнять функции нервных центров и в них могут замыкаться периферические рефлексы.
Нервная регуляция висцеральных функций может происходить в этом случае полностью в автономном режиме (отсюда название "автономная нервная система" согласно действующей Международной анатомической номенклатуре) без участия нейронов ЦНС. Передача возбуждения в вегетативных ганглиях и микроганглиях имеет свои особенности: а) выраженный феномен мультипликации (дивергенции); б) большая длительность синаптической задержки от 1,5 до 30 мс; в) низкая лабильность ганглионарных нейронов с частотой генерации потенциалов действия в среднем от 2 до 15 Гц.
Активность АНС не находится под прямым контролем сознания — это еще один признак ее автономности. Соматическая же нервная система во многих случаях контролируется сознанием.
АНС отличается от соматической нервной системы также локализацией преганглионарных нейронов и очаговым выходом их аксонов (преганглионарных волокон) из ЦНС; широким распространением в организме вегетативных нервов; малым диаметром нервных вегетативных эфферентных волокон (тип В) и отсутствием у части из них (волокон типа С) миелиновой оболочки. Поэтому вегетативные нервные волокна менее возбудимы и лабильны, длительность потенциала действия в них составляет 5—7 мс, а скорость его проведения — от 0,5 до 18 м/с.
Характерной функциональной особенностью АНС является наличие тонуса вегетативных центров — постоянной активности многих преганглионарных и ганглионарных нейронов. Формирование тонуса связано: а) со способностью клеток-осцилляторов вегетативных ганглиев и многих нейронов вегетативных центров ЦНС к спонтанной активности (автоматии); б) постоянным поступлением к ним информации от периферических сенсорных рецепторов; в) действием на них биологически активных веществ и метаболитов.
Значение тонуса вегетативных центров заключается в создании постоянного контроля над клетками, обладающими собственной активностью, и обеспечении их возможности к ответу двойной направленности (активации или торможению функции), а также в коррекции активности иннервируемых клеток в плавном режиме и без промедления.
автономный рефлекс
Механорецепторы, хеморецепторы, ноцирецепторы, терморецепторы и полимодальные рецепторы сосудов и внутренних
2 органов
Нейрон спинномозгового узла
Боковые
соматическим
рефлекс
Рецепторы
мышц, сухожилий, суставов, кожи
Задние
рога
Экстра-
или интрафуэальные мышечные волокна
скелетной мышцы
Преганглио-
Постганглио- нарное нарное волокно
волокно
Рис.
7.14.Схема
рефлекторных дуг соматического и
автономного (вегетативного) рефлексов.
Звенья рефлекторной дуги: / - рецепторное;
2— афферентное;
3— центральное
(вставочное);
4 -эфферентное;
5 — исполнительное (рабочий орган,
эффектор); АХ — ацетилхолин; Н-ХР —
никотиновый холинорецептор
постсинаптической мембраны (Н-ХРМТ— Н-ХР мышечного типа; Н-ХРн- Н-ХР
нейронального типа); М-ХР — муска-
риновый холинорецептор постсинаптической
мембраны (М|, М2, М3—подтипы ХР);
НА - норадреналин;
а-,р-АР —
альфа-, бета-адренорецепторы
постсинаптической мембраны
Спинной
мозг
Особенности рефлекторной дуги АНС представлены на рис. 7.14. Дуга центрального вегетативного рефлекса включает как минимум четыре нейрона: чувствительный (в спинномозговом ганглии), промежуточный, преганглионарный (в спинном мозге) и эфферентный (в вегетативном ганглии). Одна из главных особенностей периферического вегетативного рефлекса заключается в том, что его дуга может замыкаться вне ЦНС в вегетативных ганглиях и состоять в самом простом варианте только из двух вегетативных нейронов — афферентного и эфферентного. Такие рефлексы называют периферическими.
Классификации вегетативных рефлексов. Вегетативные рефлексы подразделяют:
по уровню замыкания рефлекторной дуги: центральные (спинномозговые, стволовые, гипоталамические, корковые) и периферические (интра- и экстраорганные, а также аксон-рефлексы);
по расположению рецепторов и органа эффектора:
висцеро-висцеральные рефлексы, вызывающие изменение деятельности внутренних органов в ответ на раздражение сенсорных рецепторов этих же или других висцеральных органов и/или сосудов;
сомато-висцеральные рефлексы, вызывающие изменение активности внутренних органов в ответ на раздражение сенсорных рецепторов скелетных мышц, суставов, кожи, органов чувств. Афферентное звено этих рефлексов относится к соматической нервной системе, а эфферентное — к АНС. Примером может служить глазосердечный рефлекс Дани- ни — Ашнера — уменьшение частоты сердечных сокращений (пульса) при надавливании на глазные яблоки. Сомато- висцеральные рефлексы лежат в основе вегетативного обеспечения физической и психической деятельности человека. Так, например, раздражение проприорецепторов мышц во время физической работы ведет к усилению дыхания, сердечной деятельности, потоотделения;
висцеро-соматические рефлексы, вызывающие изменение соматической деятельности при возбуждении сенсорных рецепторов внутренних органов или сосудов. Проявления вис- церо-соматических рефлексов могут служить важным диагностическим признаком. Так, например, при аппендиците и холецистите над очагом поражения имеет место защитное напряжение мышц. Оно обусловлено возбуждающим действием чувствительных вегетативных афферентных волокон от пораженного органа на мотонейроны соответствующих сегментов спинного мозга.
Нейрохимические особенности. Передача информации между нейронами в ЦНС и в вегетативных ганглиях, а также между нейронами и иннервируемыми клетками других тканей осуществляется через синапсы тремя способами: электрическим, химическим и смешанным. Основным из них является химический способ с помощью веществ, называемых медиаторами. Он осуществляется по определенным закономерностям согласно двум принципам. Первый заключается в том, что нейрон со всеми своими отростками выделяет один основной медиатор (1-й принцип Дейла) и, как установлено в последние десятилетия, нескольких дополнительных. Их называют ко- медиаторами или ко-трансмиттерами. Второй принцип гласит, что действие каждого медиатора на клетку-мишень (нейрон, миоцит, железистую клетку) зависит от природы рецептора постсинаптической мембраны.
Медиатором преганглионарных нервных волокон во всех вегетативных ганглиях и мозговом веществе надпочечников является АХ. Он действует на ганглионарные нейроны через никотиновые холинорецепторы (Н-ХР), называемые так из-за их высокой чувствительности к АХ и никотину. Доказано наличие двух подтипов Н-ХР: нейронального (Н-ХРн) и мышечного (Н-ХРМТ). Таким образом, все выходящие из ЦНС нервные волокна имеют в качестве основного медиатора АХ, который действует на эффекторные клетки посредством активации их Н-ХР
Медиаторами постганглионарных нервных волокон в синапсах(нейро- эффекторных соединениях) на гладких миоцитах и других клетках могут быть АХ, норадреналин (НА) и другие химические вещества (дофамин, се- ротонин, гистамин, АТФ). Основными медиаторами являются АХ и НА. В этом случае АХ действует на клетки через особый тип рецепторов — мус- кариновые ХР (М-ХР), которые высокочувствительны также и к алкалоиду мускарину, но не к никотину. Их стимуляция АХ предупреждается (блокируется) веществами типа атропина, получившими название М-холи- ноблокаторов. Доказано наличие трех подтипов М-ХР(М]. 2- 3..ХР).
Отделы АНС и их функции. В АНС выделяют два главных отдела — симпатический и парасимпатический. Важное место в АНС отводят также метасимпатическому отделу.
Симпатический (тораколюмбальный, адренерги- ческий) отдел АНС. Симпатические преганглионарные нейроны расположены в вегетативных ядрах боковых рогов серого вещества сегментов спинного мозга, начиная с 8-го шейного, всех грудных и заканчиваясь во 2-м или в 3-м поясничном сегменте включительно (отсюда анатомическое название этого отдела — тораколюмбальный). Из спинного мозга белые преганглионарные симпатические волокна (типа В) выходят в составе передних корешков и, отделившись от них, идут к симпатическим стволам (параверте- бральным ганглиям), расположенным по обе стороны позвоночника. Часть волокон прерывается в этих ганглиях, а часть — проходит транзитом. Поэтому из каждого ганглия симпатического ствола выходит два типа вегетативных нервных волокон. Первый из них — серые постганглионар- ные безмиелиновые волокна (типа С), идущие непосредственно к клеткам иннервируемых вегетативных органов (сердцу, слюнным железам, сосудам ЦНС), а также в составе соматических нервов к коже, скелетным мышцам, потовым железам, пиломоторным мышцам. Второй тип — это преганглионарные волокна, идущие в периферические (превертебральные и концевые) ганглии, а также в мозговое вещество надпочечников. Серые постганглионарные волокна (типа С) иннервируют органы брюшной полости, малого таза и наружные половые органы.
Выделяют три варианта симпатической иннервации органов: прямой контакт с эффекторными клетками через синапс и непрямой контакт через иннервацию сосудов органа (и уже от них — диффузия медиатора к клеткам органа) или через взаимодействие с другими отделами АНС. Симпатические волокна иннервируют большинство органов и тканей организма, включая ЦНС и скелетные мышцы.
Симпатические ганглии обычно удалены от иннервируемых ими органов. Поэтому преганглионарные волокна обычно короткие или средней длины, а постганглионарные — длинные или средние. В симпатических ганглиях выражено явление мультипликации (дивергенции). Одно преганглионарное волокно передает сигнал ко многим ганглионарным нейронам. Постганглионарные симпатические волокна могут ветвиться и образовывать целые сплетения в иннервируемых органах, контактировать и передавать информацию сразу сотням или тысячам эффекторных клеток за счет выделения медиатора не только из нервных окончаний, но и из расширений (варикозов) аксона. За счет этого влияние симпатического отдела АНС приобретает генерализованный характер.
Медиатором преганглионарных симпатических волокон в ганглиях является АХ (рис 7.14), который действует через Н-ХРн. Постганглионарные симпатические волокна (90% из них) выделяют норадреналин (НА) (отсюда название отдела — адренергический), который действует на эффекторные клетки через а- и р-АР. Вместе с НА могут выделяться ко-медиаторы (нейропелтид Y, АТФ), ферменты и другие субстанции. Из симпатических волокон в потовых железах выделяется АХ (действует через М3-ХР), а в сосудах почек — дофамин (действует через Dj- и 02-дофаминовые рецепторы).
При повышении тонуса симпатического отдела АНС происходит: расширение зрачков (мидриаз), сокращение гладких мышц сфинктеров и расслабление гладких мышц полых органов — трахеи и бронхов и улучшение их проходимости, расслабление желудка и кишечника и торможение их перистальтики, расслабление мочевого пузыря и накопление в нем мочи, повышение активности кардиомиоцитов — увеличение частоты и силы сокращения сердца.
Под влиянием повышения активности симпатических нервных волокон происходит также стимуляция гликогенолиза (распада гликогена) и гликолиза в скелетных мышцах и повышение их работоспособности; активация образования глюкозы из других органических веществ — жирных кислот и аминокислот в печени и повышение уровня глюкозы в крови, а также стимуляция липолиза в жировой ткани и повышение уровня липидов в крови; стимуляция секреции нейрогормонов хро- маффинными клетками мозгового вещества надпочечников.
Среди эффектов повышения тонуса симпатических нейронов следует отметить: сужение артериальных сосудов органов брюшной полости, кожи, слизистых оболочек и большинства вен; увеличение потоотделения; уменьшение секреции желез желудочно-кишечного тракта; повышение возбудимости сенсорных рецепторов и нейронов ЦНС.
Симпатический отдел АЦС способствует повышению работоспособности организма, активирует метаболизм тканей, что получило название адаптационно-трофического влияния.
Симпатический отдел выполняет эрготропную функцию и тем самым обеспечивает мобилизацию резервов организма и возможность его активного взаимодействия с факторами внешней среды. Именно поэтому возбуждение симпатического отдела имеет место всякий раз в период стресса (реакций организма, направленных на выживание в экстремальных ситуациях).
В свою очередь симпатический (тораколюмбальный) отдел АНС контролируется и находится под регулирующими влияниями высших вегетативных центров гипоталамуса, мозжечка, ствола, подкорковых структур и коры полушарий большого мозга.
Парасимпатический (краниосакральный, холинер- гический) отдел АНС. Он выполняет три основные функции: 1) передает в ЦНС информацию от сенсорных рецепторов сосудов и внутренних органов; 2) снабжает моторными и секреторными волокнами гладкую мускулатуру, железы, сердце и внутренние органы; 3) оказывает трофотропное действие и способствует восстановлению нарушенного гомеостаза.
Центральные парасимпатические преганглионарные нейроны расположены на большом удалении друг от друга в трех частях ЦНС: среднем мозге, продолговатом мозге и в сакральном отделе спинного мозга (отсюда анатомическое название этого отдела — краниосакральный).
Афферентные парасимпатические волокна передают в ЦНС информацию от механо-, хемо-, термо- и полимодальных сенсорных рецепторов сосудов и внутренних органов о параметрах гомеостатических показателей внутренней среды. Наиболее важными афферентными парасимпатическими волокнами являются волокна, входящие в состав языкоглоточного (50%) и особенно блуждающего (до 90%) нервов. В блуждающем нерве имеется множество афферентных волокон от сенсорных рецепторов сосудов, легких, сердца, органов брюшной полости.
Эфферентные парасимпатические волокна делят на три группы в зависимости от места их выхода из ЦНС.
Исходящие из среднего мозга в составе глазодвигательного нерва (III пара черепных нервов) для иннервации сфинктера зрачка и ресничной мышцы глаза. При активации этих мышц наблюдается миоз (сужение зрачка) и увеличение преломляющей силы хрусталика глаза.
Исходящие из продолговатого мозга и входящие в состав лицевого (VII пара черепных нервов), языкоглоточного (IX пара) и блуждающего (X пара) нервов. Парасимпатические волокна лицевого нерва снабжают подчелюстные и подъязычные слюнные железы, слезные железы и железы слизистых оболочек полостей носа, рта, неба и их сосуды, а также-гладкие мышцы сосудов мозговой оболочки секреторно-моторными и сосудорасширяющими волокнами. Парасимпатические эфферентные волокна языкоглоточного нерва идут к околоушной слюнной железе, железам оболочки щек и губ, а также зева и корня языка, усиливают секрецию. Эфферентные парасимпатические волокна блуждающего нерва идут к органам шеи, груди и брюшной полости, за исключением органов, расположенных в его нижней трети. Вызывают угнетение работы сердца, сужение бронхов, усиление моторики кишечника, увеличение секреции желудочных, кишечных, бронхиальных желез.
3. Аксоны сакрального отдела начинаются от преганглио- нарных нейронов II, III, IV крестцовых сегментов спинного мозга и идут к мочевому пузырю, прямой кишке и половым органам, снабжая их секреторно-моторными и сосудорасширяющими волокнами.
Парасимпатическая иннервация значительно менее обильная, чем симпатическая. Концевые ганглии обычно располагаются вблизи, на поверхности (экстрамурально) или внутри (интрамурально), в самом иннервируемом органе. Поэтому преганглионарные волокна — длинные, а постганглионарные — короткие. Все эфферентные парасимпатические волокна тонкие и миелинизированные, относятся к типу В. В парасимпатических ганглиях обычно одно преганглионарное волокно образует синапс с одним ганглионарным нейроном. Поэтому парасимпатическая регуляция достаточно точная и локальная. Выделяют два варианта парасимпатической иннервации органов: прямой контакт с эффекторными клетками через нейроэффек- торное соединение (синапс) и непрямой контакт через взаимодействие с другими отделами АНС.
Медиатором парасимпатических преганглионарных волокон является АХ, который действует через Н-ХРн. Из окончаний парасимпатических постганглионарных волокон выделяется также АХ (отсюда название — холинергический отдел). Вместе с АХ выделяются ко-медиаторы ВИП, АТФ. АХ действует на эффекторные клетки через М2-И М3-ХР.
Метасимпатический (интраорганный, неадренер- гический и нехолинергический) отдел АНС — это комплекс микроганглионарных образований, формирующих нервные сплетения и расположенных в стенках внутренних органов (отсюда название — интраорганный отдел). Речь вдет о наличии микроганглиев в желудке, кишечнике, мочевом пузыре, сердце, бронхах. Наиболее изучены метасимпатические отделы кишечника и сердца. Метасимпатический отдел кишечника (часто называемый энтеральной нервной системой) включает межмышечное (ауэрбахово) и подслизистое (мейснерово) сплетения. Они состоят из множества микроганглиев, к которым передаются импульсации от симпатического и парасимпатического отделов АНС, а также от собственных афферентных нейронов, тела которых находятся в микроганглиях и передают информацию от сенсорных рецепторов стенки кишечника.
Постганглионарные волокна эфферентных нейронов мета- симпатической системы идут к миоцитам и железистым клеткам кишечника и контролируют их активность. Центральные симпатические и парасимпатические влияния играют, по-видимому, модулирующую роль. Потери входа импульсации от этих отделов АНС (например, при операциях пересадки кишечника) не уменьшают активности нейронов в пересаженных сплетениях. Эта активность обеспечивает саморегуляцию функций кишечника. Метасимпатический отдел обладает большей независимостью от ЦНС, чем другие отделы АНС.
В метасимпатическом отделе АНС наряду с холинергически- ми и адренергическими нейронами широко представлены нехо- линергические и неадренергические нейроны (отсюда название - нехолинергический и неадренергический отдел). В качестве медиаторов в них могут находиться пептиды (динорфин, холецис- токинин, нейропептид Y, соматостатин, ВИП), биогенные амины (серотонин, гистамин, мелатонин), пурины (АТФ) и др. Некоторые моторные нейроны содержат одновременно до пяти различных медиаторов. Вероятно, что это необходимо для очень тонкой регуляции работы эффекторных клеток (например, для управления моторикой кишечника, приуроченной к процессам гидролиза и всасывания питательных веществ).
Выделяют три основные функции метасимпатического отдела АНС:
выполняет роль периферических нервных центров и обеспечивает постоянный и непрерывный контроль за работой внутренних органов;
участвует в поддержании гомеостаза;
участвует в передаче информации от сенсорных рецепторов внутренних органов к ЦНС.
Взаимодействие отделов АНС в регуляции вегетативных функций. Реализуется на двух уровнях: периферическом и центральном.
Взаимодействие на периферическом уровне имеет место на эффекторных клетках, которые получают двойную (или даже тройную) вегетативную иннервацию. Основой этого взаимодействия является антагонизм во влиянии на иннервируемые клетки парасимпатического и симпатического отделов АНС- Так, стимуляция симпатических нервов вызывает усиление сердечной деятельности, торможение перистальтики кишечника, а парасимпатических волокон вагуса — угнетение работы сердца, стимуляцию моторики кишечника. Антагонистические эффекты взаимодействия имеют место и на других уровнях. Передача возбуждения в симпатических ганглиях тормозится парасимпатическими влияниями, а в парасимпатических ганглиях — симпатическими.
Взаимодействие различных отделов АНС на центральном уровне носит сложный, кооперативный и взаимоконтролирую- щий характер. Функции преганглионарных нейронов АНС контролируются высшими вегетативными надсегментарными центрами, расположенными в различных отделах головного мозга: 1) стволе мозга — сосудодвигательный центр, голубое пятно, ядра шва, центр рвоты; 2) мозжечке; 3) промежуточном мозге — гипоталамус, таламус; 4) концевом мозге — базальные ганглии, кора больших полушарий.
Надсегментарные образования обеспечивают осуществление инте- гративных функций при организации целесообразной адаптивной деятельности. Надсегментарные механизмы регуляции вегетативных функций имеют три важные особенности: I) в них отсутствуют специфические вегетативные нейроны; 2) поражения надсегментарных центров могут проявляться не только вегетативными расстройствами, но и нарушением соматических (психических и/или двигательных) функций; 3) надсегментарные центры используют для организации адаптивного взаимодействия всех отделов АНС. Особое место винтеграции работы различных отделов АНС отводят гипоталамусу.
Вегетативное обеспечение различных форм физической и психической деятельности организма. Реакция организма на любой вид нагрузки включает в себя интенсификацию энергетического обмена. А это требует увеличения доставки дополнительного количества кислорода и питательных веществ к работающим органам. В связи с этим при любой нагрузке всегда отмечается активизация сердечной деятельности (увеличение силы и частоты сердечных сокращений), увеличение минутного объема дыхания. Увеличение теплопродукции при мышечной работе требует усиления процессов теплоотдачи, т.е. расширения сосудов кожи и усиления работы потовых желез. Таким образом, в единстве с анимальным (двигательным или умственным) динамическим стереотипом формируется вегетативный динамический стереотип, представля-
Таблица 7.1. Критерии оценки исходного вегетативного тонуса
|
Симптомы и показатели |
Симпатическая реакция |
Парасимпатическая реакция |
|
Зрачки |
Расширены |
Нормальные |
|
Температура тела |
Повышена (37 °С и выше) |
Снижена (днем ниже 36 °С) |
|
Переносимость холода |
Удовлетворительная |
Плохая |
|
Переносимость тепла |
Непереносимость жары |
Удовлетворительная |
|
Масса тела |
Склонность к похуданию |
Склонность к прибавке массы |
|
Пульс |
Тахикардия (более 90 ударов в минуту) |
Брадикардия (менее 60 ударов в минуту) |
|
Артериальное давление |
Повышено |
Понижено или норма |
|
Дыхание |
Нормальное или учащенное |
Медленное и глубокое |
|
Работоспособность |
Повышена |
Снижена |
|
Число эритроцитов в крови |
Увеличено |
Уменьшено |
|
Лейкоциты и нейтро- филы |
Увеличенное количество |
Уменьшенное количество |
|
Лимфоциты и эози- нофилы |
Нормальное количество |
Увеличенное количество |
ющий собой комплекс условных и безусловных рефлексов, обеспечивающих регуляцию вегетативных функций в соответствии с уровнем физической и/или умственной активности.
Объективные и субъективные показатели оценки функционального состояния различных отделов АНС.
Выделяют три группы показателей:
1-я группа — показатели исходного вегетативного тонуса. Они выборочно изложены в табл. 7.1.
2-я группа показатели вегетативной реактивности. Вегетативные реакции, возникающие в ответ на действие внешних и внутренних раздражителей, характеризуют вегетативную реактивность. При этом существенна сила реакции (размах колебаний вегетативных показателей) и ее длительность (время возврата вегетативных показателей к исходному уровню). При исследовании вегетативной реактивности необходимо учитывать закон исходного уровня. Согласно ему, чем выше исходный уровень, тем в более деятельном и напряженном состоянии находится система или орган, тем меньший ответ возможен при действии возмущающих стимулов (вплоть до парадоксальной реакции).
Выделяют две группы методов исследования вегетативной реактивности: 1) фармакологические методы — введение нор- адреналина или ацетилхолина, их агонистов (веществ, действующих подобно этим медиатором) или антагонистов (веществ, блокирующих действие медиаторов); 2) физические методы — холодовая или тепловая проба, изменение диаметра зрачка (при закрытии или открытии глаза), воздействие давлением на рефлексогенные зоны. Например, вызывают глазо- сердечный рефлекс Данини — Ашнера — замедление сердечной деятельности при надавливании на глазные яблоки из-за повышения тонуса вагуса, а также солярный, или эпигаст- ральный рефлекс Тома и Ру — замедление частоты сердечных сокращений при надавливании на солнечное сплетение до ощущения пульсации брюшной аорты.
• 3-я группа показатели вегетативного обеспечения деятельности организма. В клинической физиологии оно проводится с помощью экспериментального моделирования разных видов деятельности: физической (дозированная физическая нагрузка), умственной (счет в уме), эмоциональной (моделирование положительных и отрицательных эмоций).
7.12. Цереброспинальная жидкость
Цереброспинальная жидкость (ЦСЖ) заполняет систему желудочков, каналов и субарахноидальное пространство головного и спинного мозга. Цереброспинальную жидкость также называют ликвором и говорят о ликворных пространствах мозга. У взрослого человека в ликворных пространствах находится около 140 мл ЦСЖ, это составляет около 10% от массы мозга. Из этого количества 25—35 мл находится в системе мозговых желудочков, примерно столько же в поясничном расширении подпаутинного пространства — концевой цистерне, из которой у человека берут пробы ЦСЖ путем пункции между 3-м и 4-м поясничными позвонками. Остальные 70— 90 мл заполняют субарахноидальные пространства и цистерны головного мозга. ЦСЖ отделена от ткани мозга слоем выстилки, называемой эпендимой. В ее структуру входит слой эпителиоподобных клеток, под которым располагается сеть глиальных клеток и волокон.
Основное количество ЦСЖ вырабатывается за счет ее секреции сосудистыми сплетениями желудочков головного мозга. Небольшое количество жидкости, видимо, может поступать в ликворные пространства и от эпендимы, выстилающей межжелудочковые участки ликворных пространств, а также путем выхода из межклеточных пространств мозга.
У человека за сутки образуется около 500 мл ликвора, происходит непрерывное его обновление (при полной замене — около 5 раз в сутки). Образующийся ликвор движется в направлении от боковых желудочков к третьему, затем четвертому желудочку. От последнего, через боковые и срединное отверстия, ЦСЖ поступает в субарахноидальное пространство головного и спинного мозга.
Из субарахноидального пространства ликвор через ворсинчатые выпячивания паутинной оболочки (пахионовы грануляции) проникает в венозную систему мозга. Затруднения циркуляции ликвора и особенно его оттока в кровь создает опасность нарушения функций центральной нервной системы.
Ликвор выполняет: 1) защитную функцию, окружая мозг и играя роль гидравлической подушки, защищающей мозг при ускорениях и ударах; 2) дренажную функцию за счет удаления жидкости и веществ, которые проникают в него из крови через гематоэнцефалический барьер и выходят из мозга с током межклеточной жидкости; 3) регуляторную функцию за счет влияния поступающих в него биологически активных веществ и метаболитов на прилежащие к ликворным пространствам структуры. Например, обнаружена важная роль изменений содержания в ликворе СО2- и Н+ ионов в регуляции дыхания.
Исследование ликвора в клинической практике проводится для диагностики ряда заболеваний. В норме ликвор — прозрачная бесцветная жидкость, состав которой отличается от такового плазмы крови прежде всего по содержанию белка и некоторых минеральных ионов (Са2+, К+). Содержание белка в ликворе (0,2 г/л) приблизительно в 40 раз меньше, чем в крови. При этом отношение глобулинов к альбуминам составляет 0,2—0,3; содержание Са2+ (1,2 мМ/л) в 2 раза, а содержание К+ (2,9 мМ/л) на 40% меньше, чем в крови. Уровень натрия в крови и ликворе приблизительно одинаков, а рН составляет соответственно 7,40
и 7,31. Снижение рН ликвора связано с относительно большим (около 50 мм рт.ст.) напряжением С02 в ликворе.
Содержание лейкоцитов в ликворе составляет в норме 1,5 млн/л. Около 60% из них представлены лимфоцитами.
Контрольные вопросы и задания
Что называют нервным центром? Приведите примеры нервных центров и их функций.
Перечислите методы исследования функций нервных центров.
Что такое электроэнцефалография, ЭЭГ-ритмы? Дайте им характеристику.
Каковы функции, центры и нейроны спинного мозга?
Что такое миотатические рефлексы? Особенность их рефлекторной дуги и использование в медицине.
Какова структура рефлекторной дуги оборонительных сги- бательных и перекрестныхразгибательныхрефлексов?
7 Как происходит регуляция тонуса мышц и спинальных а-мо- тонейронов за счет интеграции афферентных и эфферентных импульсов?
Что такое спинальный шок, каковы его проявления?
В чем заключается проводниковая функция спинного мозга?
Дайте характеристику продолговатого мозга и моста, их функций.
Перечислите центры продолговатого мозга и моста.
Дайте характеристику среднего мозга, его структуры и функций.
Каковы функции красного ядра и черной субстанции? Что такое децеребрационнаяригидность?
В чем заключаются статические и статокинетическиерефлексы? Каковы проявления нистагма?
Каковы функции мозжечка?
Каковы функции ретикулярной формации ствола мозга?
17 Перечислите ядра и функции таламуса.
Дайте характеристику гипоталамуса, его роли в регуляции гомеостаза, вегетативных функций, поведенческих реакций.
Каков механизм участия гипоталамуса в регуляции активности эндокринных желез и стресс-реакции?
Каковы функции базальных ганглиев?
Дайте характеристику лимбической системы и ее функций.
Охарактеризуйте кору большого мозга, ее нейронные слои, волокна, нейронную организацию рецепторных полей.
Как сенсорные области выделяют в коре полушарий большого мозга?
Охарактеризуйте топографию и функции соматосенсорной области коры.
Каковы моторные и премоторные области коры, их функции?
Перечислите пути передачи влияний коры на скелетные мышцы.
27 Каковы функции ассоциативных областей коры?
Охарактеризуйте речевые центры коры, их локализацию и функции.
Что такое ликвор, его образование и циркуляция?
Какова физиологическая роль ликвора?
Назовите состав ликвора. От чего он зависит?
Дайте определение понятий "автономная нервная система" и "соматическая нервная система"
Назовите отделы автономной нервной системы (АНС) и их функции.
Объясните, почему симпатический отдел АНС получил названия "тораколюмбальный" и "адренергический"
Объясните, почему парасимпатический отдел АНС получил названия "краниосакральный" и "холинергический"
Объясните, почему метасимпатический отдел АНС получил названия "интраорганный" и "нехолинергический, неадренерги- ческий"
37 Перечислите основные морфофункциональные и нейрохимические особенности АНС.
Назовите основные медиаторы синапсов АНС в вегетативных ганглиях и на иннервируемых эффекторных клетках. Укажите, через какие типы рецепторов действуют эти медиаторы.
Дайте определение понятия "тонус вегетативных центров" Укажите механизмы его формирования и его значение.
Назовите виды вегетативных рефлексов, приведите примеры.
Назовите морфофункциональные и нейрохимические особенности симпатического и парасимпатического отделов АНС.
Перечислите физиологические эффекты влияния симпатических и парасимпатических нервов на иннервируемые органы (сердце, железы, гладкие мышцы сфинктеров, полых органов, сосудов).
Назовите функции афферентных вегетативных нервных волокон.
Назовите основные типы нейронов вегетативных ганглиев.
Кратко опишите, как вы понимаете взаимодействие отделов АНС в регуляции вегетативных функций.
Укажите особенности надсегментарных механизмов регуляции вегетативных функций.
47 Назовите три группы показателей, позволяющих оценить функциональное состояние различных отделов АНС.
Ситуационные задачи
Зарисуйте, как отражается на ЭЭГ реакция десинхронизации. О чем она свидетельствует?
После травмы спинного мозга у больного полностью выпала чувствительность на передней поверхности бедра. Сколько сегментов повреждено?
В какой области надо раздражать кору больших полушарий, чтобы проверить наличие проведения возбуждения от коры до мышц левой руки? от коры до мышц левой ноги?
Латентный период рефлекса сгибания лапки лягушки, вызываемого раздражением кожи кислотой, составил 8 с. Как изменится латентный период этого рефлекса, если такое же раздражение наносить на фоне стимуляции гигантоклеточного ядра ретикулярной формации ствола мозга?
После травмы спинного мозга у пострадавшего полностью исчезла двигательная активность левой ноги при сохранении движений правой ноги, однако на ней понизилась тактильная и полностью выпали температурная и болевая чувствительность. На каком уровне и в какой степени нарушена целостность спинного мозга?
Нарисуйте схему рефлекторных дуг соматического и автономного рефлексов, замыкающихся в спинном мозге. На схеме укажите названия сенсорных рецепторов, места расположения эфферентных нейронов, названия эффекторных клеток.