

системные механизмы
.pdf91
30
25
20
15
%
10
5
0
-5
int |
si |
группы опытов-ритмы ЭКоГ
B2A  B1A
B1A 
 AA
AA 
 TA
TA  DA
DA
А
%
40
35
30
25
20
15
10
5
0
-5
-10
int_desin |
int_sin |
si_des |
si_sin |
|
группы опытов - ритмы ЭКоГ |
|
|
B2A  B1A
B1A 
 AA
AA 
 TA
TA  DA
DA
Б
Рисунок 3.3. Показатели ФМПА амплитуд ритмов ЭКоГ. Обозначения:А - ФМПА амплитуд ритмов ЭКоГ без использования сегментации, Б - ФМПА амплитуд ритмов ЭКоГ с использованием сегментации, B2A - ФМПА амплитуд бета-2 ритма, B1A – ФМПА амплитуд бета-1 ритма, AA – ФМПА амплитуд альфа-2 ритма, TA – ФМПА амплитуд тета ритма DA - ФМПА амплитуд дельта ритма, int – интактный мозг без сегментации, si – препарат изолированного переднего мозга без сегментации; при сегментации: int_desin – интатній мозг, десинхронизация, int_sin – интактый мозг, синхронизация, si_desin – препарат изолированного переднего
мозга, десинхронизция, si_sin - препарат изолированного92 переднего мозга, синхронизция.
ФМПА частоты бета-2, бета-1 альфа и дельта римов ЭКоГ в условиях ИМ без предварительного проведения сегментации ЭКоГ выражалась отрицательными величинами в пределах (- 0,17±0,00% - -0,64±0,00%), а ФМПА частоты тета ритма – положительной величиной 0,09±0,00%. Все показатели ФМПА амплитуды ритмов ЭКоГ препарата ИПМ были положительными и варьировали от 0,74 0,001 до 16,57 0,01 %, т.е. ЭКоГ-активность по частоте, также как и по амплитуде была «латерализована» в левое полушарие.
ЭКоГ-активность по индексам длительности бета-2, бета-1, альфа и тета ритмов ИМ и ИМП выражалась положительными величинами. Эти для ИМ были от
0,06±0,00% до 10,09±0,32%, а ИПМ от - 7,89 0,04% до
20,30 0,18%. ЭКоГ-активность по индексу длительности дельта ритма для ИМ была (-0,93±0,02%), а для ИПМ – (- 6,87 0,02%)
ФМПА показателей амплитуд ЭКоГ ИМ и ИПМ при использовании процедуры сегментации. В отрезках ЭКоГ с проявлением десинхронизации ФМПА амплитуд всех ритмов ЭКоГ, отводимой от ИМ, имела положительные значения (табл. 3.2, рис. 3.3, Б), т.е. показатели были «латерализованы в левое полушарие». Значения ФМПА для различных спектральных компонентов составляли от 1.30 ± 0.01 до 6.95
±0.09 %. В сегментах синхронизации показатели ФМПА амплитуд бета-2-, бета-1-, альфа- и тета-ритмов были отрицательными («правосторонними») и варьировали от –0.85
±0.01 до –4.15 ± 0.10 %, а показатель ФМПА амплитуды дельта-ритма оказался положительным (9.44 ± 0.16 %). Таким образом, переход паттерна ЭКоГ от десинхронизации к синхронизации в условиях интактности мозга сопровождался сменой левосторонней латерализации амплитуд бета-2-, бета- 1-, альфа- и тета-ритмов правосторонней и возрастанием левосторонней латерализации амплитуды дельта-ритма.
Все показатели ФМПА амплитуд ритмов ЭКоГ, отводимой от ИПМ, были положительными, составляя для отрезков десинхронизации от 6,85 ± 0,12 до 25,33 ± 0,49 %, а для сегментов синхронизации – от 0,17 ± 0,01 до 31,47 ± 1,99
%. Переход от десинхронизации к синхронизации ЭКоГ при93 ее отведении от ИПМ сопровождался увеличением выраженности левосторонней латерализации амплитуд бета-2-, бета-1- и альфа-ритмов и уменьшением левосторонней латерализации амплитуд тета- и дельта-ритмов.
Впериоды десинхронизации ФМПА частот бете-2 и бета-1 ритмов, ЭКоГ отводимых от ИМ выражалась отрицательными величинами (-0,01±0,00%) и (-0,02±0,00%), а ФМПА частот альфа, тета и дельта ритмов – положительными величинами в пределах 0,24±0,00% – 0,71±0,00%. В период синхронизации ФМПА частот бета-2, альфа, тета и дельта ритмов определялась отрицательными величинами в пределах (-0,03±0,00% - - 2,77±0,03%), и только показатель ФМПА частоты бета-1 ритма определялся положительным –
0,38±0,00%.
Вотрезках ЭКоГ с проявлениями десинхронизации ФМПА частот бета-2, бета-1, альфа и тета ритмов на препарате ИПМ выражалась положительными величинами в пределах 1,58±0,01% - 4,25±0,03%, а ФМПА частоты дельта-ритма – отрицательной величиной – 0,28±0,00%. В отрезках синхронизации положительность ФМПА частот бета-2, бета-1
иальфа ритмов статистически значимо увеличилась и находилась в пределах 5,70±0,08% – 11,49±0,40%. Положительность показателя ФМПА частоты тета ритма уменьшилась, а отрицательность показателя ФМПА частоты дельта ритма увеличилась до -4,61±0,10%.
Показатели ФМПА индексов длительности бета-2,бета-1
идельта ритмов ЭКоГ ИМ были отрицательны в пределах (- 0,32±0,00% - -2,17±0,03%), а альфа и тета ритмов – положительны 3,26±0.06% и 1,09±0,01% соответственно. При переходе от десинхронизации к синхронизации ФМПА индексов длительностей бета-2, бета-1 и дельта ритмов инвертировала и выражалась положительными величинами 1,26±0,03% - 13,29±0,31%. Положительность ФМПА индексов длительности альфа и тета ритмов также инвертировала и эти показатели выражались отрицательными величинами –
11,97±0,45% и 11,13±0,32%.
На препарате ИПМ ФМПА индексов длительности в отрезках десинхронизации была положительна в пределах 5,32±0,1- - 18,11±0,49%, а ФМПА индекса длительности дельта
ритма – отрицательна – 12,50±0,23%. При переходе от94 десинхронизации к синхронизации положительность ФМПА бета-1, бета-1 и альфа ритма увеличилась до 28,56±2,32% - 31,62±1,62%. Положительность ФМПА индекса длительности тета ритма уменьшилась до 2,16±0,13%. Уменьшилась также и отрицательность показателя ФМПА индекса длительности дельта ритма до (-5,27±0,17%).
3.3. Взаимоотношения амплитуд ритмов ЭКоГ в условиях ИМ и ИПМ
Структурный анализ полициклического мультиграфа, описывающего связи-отношения между показателями амплитуд ритмов ЭКоГ в левом полушарии ИМ в том случае, если сегментация не производилась, выявил три положительных связи-отношения и одну отрицательную (рис.
3.4, А З.5).
Статистически значимых коэффициентов корреляции определялось 4.
После сегментации ЭКоГ ИМ в кластере десинхронизации (рис. 3.4, Б) между амплитудами ритмов было выявлено 20 положительных связей-отношений, т.е. все узлы графа показателей ритмов ЭКоГ были связаны положительными взаимно ориентированными связями. В сегментах с проявлением синхронизации были обнаружены 14 положительных связей-отношений (В).
При расчетах коэффициентов двумерной корреляции в кластерах десинхронизации и синхронизации ЭКоГ левого полушария определялось по 10 статистически значимих коэффициентов корреляции.
При структурном анализе полициклического мультиграфа, описывающего связи-отношения между показателями амплитуд ритмов ЭКоГ в правом полушарии ИМ, которые наблюдались без применения сегментации, были выявлены 10 положительных и шесть отрицательных связейотношений (рис. 3.4, Г).
Статистически значимых коэффициентов корреляции определялось 3.
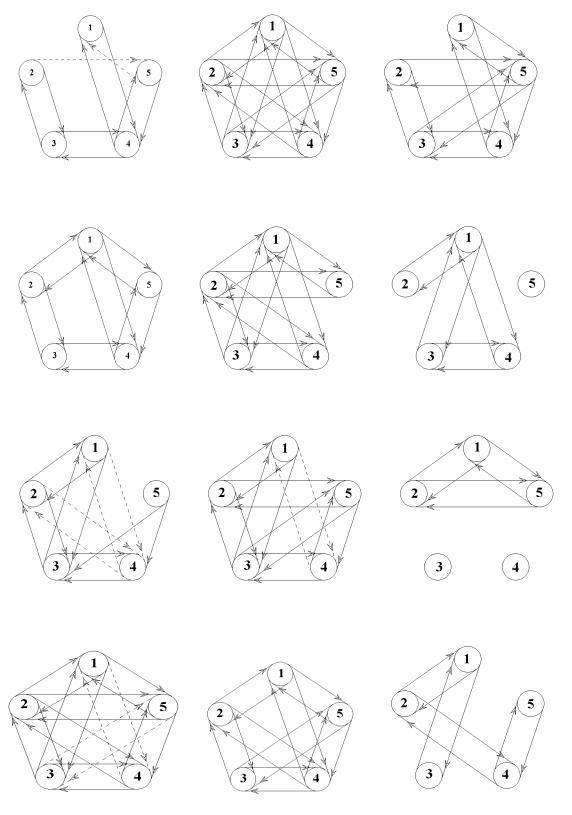
В кластере десинхронизации ЭКоГ обнаружились 1695 положительных (Д), а в кластере синхронизации – восемь положительных связей (Е).
При расчетах коэффициентов двумерной корреляции в кластерах десинхронизации и синхронизации ЭКоГ правого
А |
Б |
В |
Г |
Д |
Ж |
З |
И |
К |
Л |
М |
Рисунок 3.4. |
Математические модели |
отражающие |

взаимоотношения между амплитудами ритмов ЭКоГ левого и96 правого полушарий у интактных крыс и препарата изолированного мозга.
Обозначения: А – левое полушарие ИК без сегментации, Б – левое полушарие ИК десинхронизация, В – левое полушарие ИК, синхронизация, Г - правое полушарие ИК, без сегментации, Д - правое полушарие ИК, десинхронизация Е – правое полушарие ИК, синхронизация; Ж - левое полушарие ПМ без сегментации, З – левое полушарие ПМ десинхронизация, И – левое полушарие ПМ, синхронизация, К - правое полушарие ПМ, без сегментации, Л - правое полушарие ПМ, десинхронизация М – правое полушарие ПМ, синхронизация; 1 - амплитуда бета-2-ритма, 2 - амплитуда бета-1 ритма, 3 – амплитуда альфа ритма, 4 – амплитуда тета ритма, 5 – амплитуда дельта ритма. Сплошные линии – положительные влияния, прерывистые линии – трицательные влияния.
интактные
20 |
|
|
|
|
|
|
|
|
|
16 |
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|||
15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
л |
|
|
|
|
|
|
|
|
п |
|
4 |
|
|
4 |
|
|
|
|||
|
|
|
|
|
|
||||
5 |
|
|
3 |
|
|
|
|||
|
|
|
|
|
|
|
|
||
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
регрессии |
корреляции |
|
|
|||||
|
|
|
|
||||||
А
препарат изолированного мозга
25 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
20 |
|
|
16 |
|
|
|
|
|
14 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
л |
||
10 |
|
|
|
10 |
10 |
|
|
|
8 |
|
|
10 |
|
10 |
|
|
п |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
регрессии |
корреляции |
регрессии |
корреляции |
|
|
||||||||||||||
|
|
|
|||||||||||||||||
|
|
десинхронизация |
|
синхронизация |
|
|
|
|
|||||||||||
Б
Рисунок. 3.5. количество статистически значимых коэффициентов регрессии и двумерной корреляции между амплитудами ритмов ЭКоГ в условиях интактного мозга (А)
и препарата изолированного переднего мозга (Б) в условиях97 десинхронизации и синхронизации.
полушария определялось по 10 статистически значимих коэффициентов корреляции.
При структурном анализе полициклического мультиграфа, который описывал связи-отношения между амплитудами ритмов несегментированной ЭКоГ, отведенной от левого полушария в условиях ИПМ, выявилось 12 положительных связей-отношений и восемь отрицательных
(рис. 3.4, Ж).
При расчетах коэффициентов двумерной корреляции между амплитудами ритмов несегментированной ЭКоГ, отведенной от левого полушария в условиях ИПМ, определялось 8 статистически значимых коэффициентов корреляции.
После проведения сегментации ЭКоГ в кластере десинхронизации наблюдались 15 положительных связейотношений и две отрицательных (рис. 3.4, З), а в кластере синхронизации – лишь шесть положительных связейотношений (И).
При расчетах коэффициентов двумерной корреляции в кластерах десинхронизации определялось 9 статистически значимих коэффициента корреляции, а в кластере синхронизации – 3.
Вправом полушарии при анализе несегментированной ЭКоГ отмечались 16 положительные и четыре отрицательные связи-отношения (рис. 3.4, К). Статистически значимых коэффициентов двумерной корреляции определялось 10.
Вкластере десинхронизации определялось 16 положительных связей-отношений (Л), а в кластере синхронизации – восемь положительных (М).
При расчетах коэффициентов двумерной корреляции в кластерах десинхронизации определялось 6 статистически значимих коэффициента корреляции, а в кластере синхронизации – 5.
При отведении ЭКоГ от препарата ИПМ, который можно рассматривать как модель мозга организма, находящегося в патологическом коматозном состоянии
(Начкебия А. Я., 1989), по сравнению с условиями
интактности мозга отмечались общее снижение амплитуд98 ритмов ЭКоГ и реорганизация этих ритмов. Последняя выражалась в уменьшении индексов бета-2- и тета-ритмов и увеличении соответствующих индексов бета-1- и дельтаритмов.
Показано (В. С. Русинов, О. М. Гриндель, Г. Н. Болдырева и др., 1988), что у больных с очаговыми поражениями диэнцефальных отделов мозга спектральная характеристика ЭЭГ смещается в сторону преобладания низких частот. Данный сдвиг, по мнению авторов, свидетельствует об утрате корой в подобных условиях её ведущей роли и переходе к регуляции массовой электрической активности мозга филогенетически более старыми образованиями – в значительной степени лимбической системой.
Следует отметить, что в условиях наших опытов уменьшение индекса длительности тета-ритма и, соответственно, увеличение индекса длительности дельтаритма при отведении от препарата ИПМ по сравнению с таковым в условиях ИМ были бо лее выраженными в правом полушарии, чем в левом.
ЭКоГ, отводимая от препарата ИПМ, характеризовалась бóльшими значениями индексов длительности альфа-ритма на отрезках как десинхронизации, так и синхронизации по сравнению с картиной, наблюдаемой в условиях интактности мозга. Следует подчеркнуть, что при анализе несегментированной ЭКоГ индекс длительности альфаритма, определенный в условиях ИПМ, статистически значимо не отличался от аналогичной величины, полученной для ЭКоГ, которая отводилась от ИМ.
Переход от десинхронизации к синхронизации ЭКоГ при отведении от ИМ сопровождался сменой выраженной левосторонней латерализации амплитуд бета-2-, бета-1-, альфа- и тета-ритмов правосторонней и увеличением левосторонней латерализации амплитуды дельта-ритма. Полученные результаты в определенной степени согласуются с представлениями о возможности смены доминантности полушарий в разные фазы сна (M.
Myslobodsky, M. Mintz, B. Yedid-Levy, and V. Ben-Mayer, 1977)
и об относительном повышении активности правого
полушария во время быстрого и левого – во время медленного99
сна (L. Goldstein, N. W. Stolzfus, and J. F. Gardocki, 1972).
Переход от десинхронизации к синхронизации при отведении от препарата ИПМ сопровождался увеличением левосторонней латерализации амплитуд бета-2-, бета-1- и альфа-ритмов и уменьшением левосторонней латерализации амплитуд тета- и дельта-ритмов.
С учетом описанных выше данных можно высказать предположение, что структуры ствола (в частности, РФ) и мозжечка наряду с собственно корковыми механизмами в значительной степени обеспечивают управление ФМПА в условиях интактности мозга. Следует подчеркнуть, что показатели ФПМА в условиях ИПМ не только были положительными, т.е. проявлялась отчетливая латерализация электрической активности в левое полушарие, но и при этом их модули в два–пять раз превышали аналогичные величины, характерные для ИМ. Указанная особенность, возможно, связана с тем обстоятельством, что связи полушарий со срединными образованиями мозга разного уровня организованы различным образом. Кора левого полушария имеет более развитые связи со стволовыми структурами, тогда как правого – с диэнцефальными (Н. Н. Брагина, Т. А. Доброхотова, 1988;. Н. Болдырева, Е. В. Шарова, И. С. Добронравова, 2000). Полученные результаты также могут свидетельствовать о том, что влияния из ствола и мозжечка в некоторой степени нивелируют собственную ФМПА полушарий; такое заключение согласуется с представлениями о генерализованном преобладании ингибиторных процессов в левом полушарии, а активационных – в правом (Брагина Н. Н., Доброхотова Т. А., 1988) Болдырева Г. Н., Шарова Е. В., Добронравова И. С.,
2000)
Анализ взаимных ориентированных (векторных) влияний между показателями амплитуд ритмов ЭКоГ показывает, что значительная часть этих показателей объединена достоверными связями-отношениями, что согласуется с общими представлениями о функционировании конечного мозга как единого целого. В условиях десинхронизации ЭКоГ (рис. 3.5) выявлялось большее количество связей-отношений, чем в пределах ЭКоГ-
сегментов синхронизации. Этот результат был получен как100 для ИМ, так и для препарата ИПМ.
Согласно представлениям школы Русинова (1987), увеличение количества корреляционных связей между различными компонентами ЭЭГ и ЭКоГ отражает повышение (в целом) тонуса коры, а уменьшение – снижение этого тонуса. Связи-отношения, которыми мы в данной работе оперируем, являются коэффициентами регрессии в уравнениях множественной линейной регрессии. Мы хотели бы особо подчеркнуть, что термин «связь» в данном контексте не подразумевает наличия каких-либо морфологических связей, соединяющих те или иные структуры, а лишь отражает взаимные влияния различных компонентов ЭКоГ. Тем не менее, результаты, полученные с применением двух упомянутых статистических приемов, сходны.
Уже в первых работах М.Н. Ливанова и его сотрудников с использованием метода пространственной синхронизации биопотенциталов, было показано, что возрастание этого показателя свидетельствует о переходе к напряженной умственной деятельности (Ливанов М.Н., 1972). При прямой электростимуляции ретикулярной формации среднего мозга был получен результат, подтверждающий эти наблюдения
(Янсон З.А., 1973).
Установлено (Ливанов М. Н., Королькова Т. А., Свидерская Н. Е., 1988), что глобальная пространственная синхронизация корковых потенциалов повышается при приеме такого психостимулятора, как кофеин, а введение нейролептика клозапина приводит к противоположному эффекту.
В исследованиях Ford M.R.et all., (Ford M.R., Goethe J.W., Dekker D.K., 1986), показано, что когеренция уменьшается с возрастом, у больных параноидной шизофренией, при лечении нейролептиками, и увеличивается при лечении трициклическими антидепрессантами.
Таким образом, степень синхронизации ЭКоГактивности существенно зависит от фармакоиндуцированных активационных или ингибиторных влияний.
При анализе приходящих и отходящих влияния к узлам полициклических мультиграфов, являющихся аналогами показателей амплитуд, выявляются как положительные, так и