

2.Окислительное фосфорилирование. Пункты сопряжения окисления и фосфорилирования. Атф-синтетаза митохондрий.
Окислительное фосфорилирование — один из важнейших компонентов клеточного дыхания, приводящего к получению энергии в виде АТФ. Субстратами окислительного фосфорилирования служат продукты расщепления органических соединений — белки, жиры и углеводы.
Окислительное фосфорилирование, синтез АТФ из аденозиндифосфата и неорганического фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении орг. веществ в процессе клеточного дыхания. В общем виде окислительное фосфорилирование и его место в обмене веществ можно представить схемой:
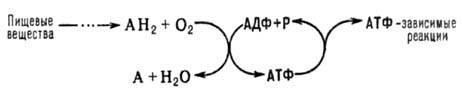
Механизм
окислительного фосфорилирования
можно представить схемой:
Перенос электронов
(дыхание)
![]() А
~ В
А
~ В![]() АТФ
А ~ В-высокоэнергетич. интермедиат
Предполагалось, что А ~ В - хим. соед. с
макроэргич. связью, напр. фосфорилир.
фермент
дыхат. цепи (хим. гипотеза сопряжения),
или напряженная конформация
к.-л. белка,
участвующего в окислительном
фосфорилировании
(конформац. гипотеза сопряжения). Однако
эти гипотезы не получили эксперим.
подтверждения. Наиб. признанием пользуется
хемиосмотич. концепция сопряжения,
предложенная в 1961 П. Митчеллом. Согласно
этой теории, своб. энергия транспорта
электронов
в дыхат. цепи затрачивается на перенос
из митохондрий
через митохондриальную мембрану
на ее наружную сторону ионов
Н+.
В результате на мембране
возникает разность электрич. потенциалов
АТФ
А ~ В-высокоэнергетич. интермедиат
Предполагалось, что А ~ В - хим. соед. с
макроэргич. связью, напр. фосфорилир.
фермент
дыхат. цепи (хим. гипотеза сопряжения),
или напряженная конформация
к.-л. белка,
участвующего в окислительном
фосфорилировании
(конформац. гипотеза сопряжения). Однако
эти гипотезы не получили эксперим.
подтверждения. Наиб. признанием пользуется
хемиосмотич. концепция сопряжения,
предложенная в 1961 П. Митчеллом. Согласно
этой теории, своб. энергия транспорта
электронов
в дыхат. цепи затрачивается на перенос
из митохондрий
через митохондриальную мембрану
на ее наружную сторону ионов
Н+.
В результате на мембране
возникает разность электрич. потенциалов
![]() и
разность хим. активностей
ионов
Н+
и
разность хим. активностей
ионов
Н+
![]() (внутри
митохондрий
рН выше, чем снаружи). В сумме эти
компоненты дают трансмембранную разность
электрохим. потенциалов ионов
водорода
(внутри
митохондрий
рН выше, чем снаружи). В сумме эти
компоненты дают трансмембранную разность
электрохим. потенциалов ионов
водорода
![]() между
матриксом митохондрий
и внеш. водной фазой, разделенными
мембраной.
Энергия
между
матриксом митохондрий
и внеш. водной фазой, разделенными
мембраной.
Энергия
![]() ,
выделяющаяся при движении протонов
внутрь митохондрий
по электрич. полю в сторону меньшей их
концентрации,
используется
АТФ-синтетазой для
синтеза
АТФ.
Т. обр., схему окислительного
фосфорилирования,
согласно этой концепции, можно представить
в след. виде:Перенос электронов
(дыхание)
,
выделяющаяся при движении протонов
внутрь митохондрий
по электрич. полю в сторону меньшей их
концентрации,
используется
АТФ-синтетазой для
синтеза
АТФ.
Т. обр., схему окислительного
фосфорилирования,
согласно этой концепции, можно представить
в след. виде:Перенос электронов
(дыхание)
![]() АТФ
АТФ
Сопряжение
окисления
и фосфорилирования
через![]() позволяет объяснить, почему окислительное
фосфорилирование,
в отличие от гликолитич. ("субстратного")
фосфорилирования,
протекающего в р-ре, возможно лишь в
замкнутых мембранных структурах, а
также почему все воздействия, снижающие
электрич. сопротивление и увеличивающие
протонную проводимость мембраны,
подавляют ("разобщают") окислительное
фосфорилирование.
Энергия
позволяет объяснить, почему окислительное
фосфорилирование,
в отличие от гликолитич. ("субстратного")
фосфорилирования,
протекающего в р-ре, возможно лишь в
замкнутых мембранных структурах, а
также почему все воздействия, снижающие
электрич. сопротивление и увеличивающие
протонную проводимость мембраны,
подавляют ("разобщают") окислительное
фосфорилирование.
Энергия
![]() ,
помимо синтеза
АТФ,
может непосредственно использоваться
клеткой
для др. целей - транспорта метаболитов,
движения (у бактерий), восстановления
нико-тинамидных коферментов
и др.
,
помимо синтеза
АТФ,
может непосредственно использоваться
клеткой
для др. целей - транспорта метаболитов,
движения (у бактерий), восстановления
нико-тинамидных коферментов
и др.
Показано, что окислительное фосфорилирование сопряжено с переносом электронов по цепи дыхательных ферментов, встроенных во внутреннюю мембрану митохондрий. Электроны поступают в дыхательную цепь от восстановленного НАД (или НАДФ) и через кофермент Q последовательно передаются от соединений с более отрицательным окислительно-восстановительным потенциалом к соединениям с более положительным потенциалом. Перенос электронов по цепи завершается восстановлением O2 с помощью цитохромоксидазы. Таким образом, процесс окисления субстрата кислородом опосредован серией окислительно-восстановительных реакций; в результате энергия, запасённая в молекуле окисляемого субстрата, освобождается небольшими порциями, что позволяет клетке использовать её более полно. Утилизация высвобождаемой энергии происходит в так называемых пунктах энергетического сопряжения. Синтез АТФ осуществляется ферментным комплексом — АТФ-синтетазой, который может катализировать и обратную реакцию. Эффективность окислительного фосфорилирования оценивают с помощью коэффициента фосфорилирования Фн/O — отношения числа молекул неорганического фосфата, связанного при фосфорилировании АДФ, к поглощённому O2; величина этого коэффициента зависит от окисляемого субстрата, физиологического состояния клеток и состава окружающей клеток среды.
Упрощённая схема цепи дыхательных ферментов, локализованных в митохондриях. Перенос электронов по цепи на трёх этапах (так называемым пунктах сопряжения) сопровождается запасанием выделяющейся энергии в форме электрохимического градиента ионов водорода, далее энергия расходуется для синтеза АТФ.

Работа АТФ-синтазы
Н+-транслоцирующая АТФ-синтаза состоит из двух частей: встроенного в мембрану протонного канала (F0) из по меньшей мере 13 субъединиц и каталитической субъединицы (F1), выступающей в матрикс. «Головка» каталитической части образована тремя α- и тремя β-субъединицами, между которыми расположены три активных центра. "Ствол" структуры образуют полипептиды F0-части и γ-, δ- и ε-субъединиц головки.
Каталитический цикл подразделяется на три фазы, каждая из которых проходит поочередно в трех активных центрах. Вначале идет связывание АДФ (ADP) и Ρi (1), затем образуется фосфоангидридная связь (2) и, наконец, освобождается конечный продукт реакции (3). При каждом переносе протона через белковый канал F0 в матрикс все три активных центра катализируют очередную стадию реакции. Предполагается, что энергия протонного транспорта прежде всего расходуется на поворот γ-субъединицы, в результате которого циклически изменяются конформации α- и β-субъединиц.
Процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — Протонная АТФ-синтаза, состоящая из 9 субъединиц 5 типов: