

Билет №25 Репликация днк
Реплика́ция ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. По имеющимся данным, в репликации ДНК, включающей узнавание точки начала процесса, расплетение родительских цепей ДНК в репликационной вилке, инициацию биосинтеза дочерних цепей и дальнейшую их элонгацию и, наконец, окончание (терминация) процесса, участвует более 40 ферментов и белковых факторов, объединенных в единую ДНК-репликазную систему, называемую реплисомой.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Ферменты хеликазы (расплетают концы), топоизомеразы (раскручивают суперспирализованные витки) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимераз, способных распознать и исправить ошибку. Источниками энергии и одновременно с этим субстратами являются dATP, dGTP, dTTP, dCTP. У прокариот выделено три типа ДНК-полимераз. Функцию элонгации выполняет ДНК-полимераза ІІІ. ДНК-полимеразы І и ІІ выполняют репарационные функции. Для затравки (инициации) требуется олигорибонуклеотид, который синтезируется праймазой.
Свойства процесса репликации:
матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности;
полуконсервативный — одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая — материнской;
идёт в направлении от 5’-конца новой молекулы к 3’-концу;
полунепрерывный — одна из цепей ДНК синтезируется непрерывно, а вторая — в виде набора отдельных коротких фрагментов (фрагментов Оказаки);
начинается с определённых участков ДНК, которые называются сайтами инициации репликации (англ. origin).
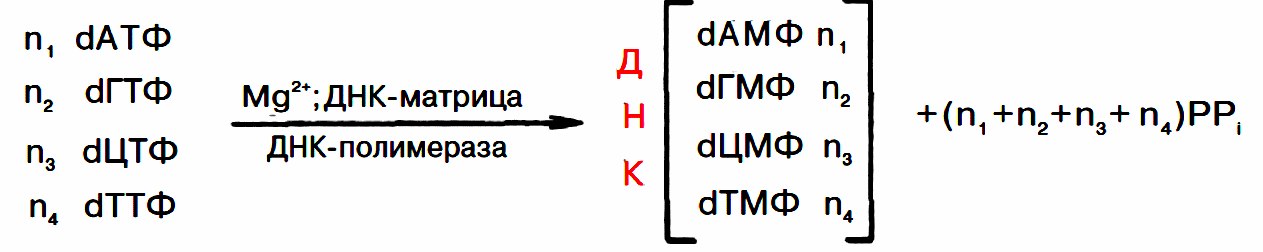
Сложность процесса репликации ДНК объясняется тем, что обе цепи реплицируются одновременно, хотя имеют разное направление (5'–>3' и 3'–>5'); кроме того, рост дочерних цепей также должен происходить в противоположных направлениях. Элонгация каждой дочерней цепи может осуществляться только в направлении 5'–>3'. Р. Оказаки высказал предположение, подтвержденное экспериментальными данными, что синтез одной из дочерних цепей осуществляется непрерывно в одном направлении, в то время как синтез другой дочерней цепи происходит прерывисто, путем соединения коротких фрагментов (в честь автора названы фрагментами Оказаки), в свою очередь синтезирующихся в противоположном направлении.
Как видно, синтез ведущей цепи ДНК идет всегда в направлении 5'–>3', соответствующем направлению движения репликационной вилки. Сохраняя правило синтеза дочерних молекул ДНК 5'–>3', синтез на второй цепи родительской ДНК идет в направлении, противоположном движению репликационной вилки. В зависимости от типа клетки фрагменты Оказаки имеют разные размеры – от нескольких сот до нескольких тысяч нуклеотидов (150–200 у эукариот и 1000–2000 у бактерий).
О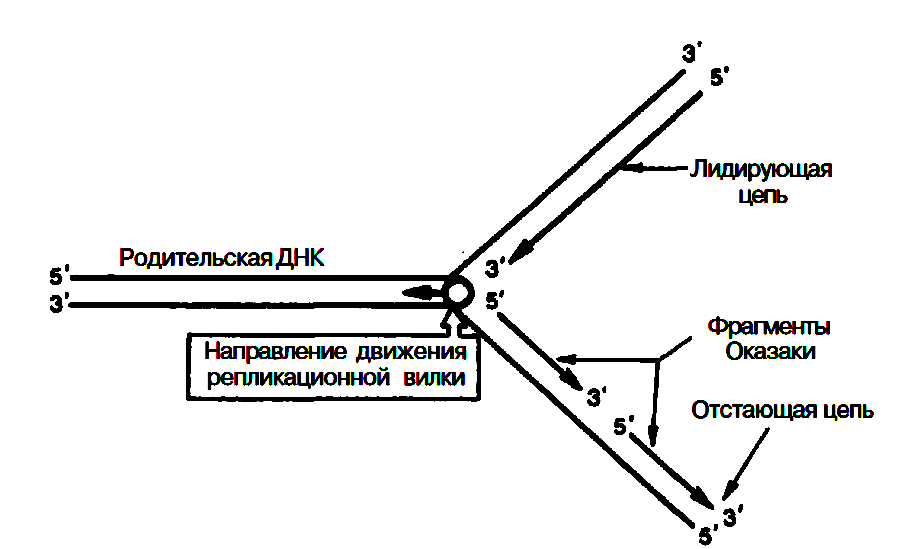 бразование
каждого фрагмента Оказаки требует
наличия короткого затравочного
комплементарного праймера – участка
РНК, синтез которого катализируется
праймазой. Затем при участии ДНК-полимеразы
III синтезируются длинные участки ДНК.
РНК-затравки далее вырезаются при
участии ДНК-полимеразы I, а свободные
места их (бреши) замещаются (достраиваются)
комплементарными дезоксирибонуклеотидами
под действием той же ДНК-полимеразы I;
наконец, сшивание разъединенных участков
отстающей цепи осуществляется при
помощи ДНК-лигаз.
бразование
каждого фрагмента Оказаки требует
наличия короткого затравочного
комплементарного праймера – участка
РНК, синтез которого катализируется
праймазой. Затем при участии ДНК-полимеразы
III синтезируются длинные участки ДНК.
РНК-затравки далее вырезаются при
участии ДНК-полимеразы I, а свободные
места их (бреши) замещаются (достраиваются)
комплементарными дезоксирибонуклеотидами
под действием той же ДНК-полимеразы I;
наконец, сшивание разъединенных участков
отстающей цепи осуществляется при
помощи ДНК-лигаз.
По имеющимся данным, в репликации ДНК, включающей узнавание точки начала процесса, расплетение родительских цепей ДНК в репликационной вилке, инициацию биосинтеза дочерних цепей и дальнейшую их элонгацию и, наконец, окончание (терминация) процесса, участвует более 40 ферментов и белковых факторов, объединенных в единую ДНК-репликазную систему, называемую реплисомой. Рассмотрим подробнее её компоненты в прокариотической клетке:
Основным ферментом, катализирующим биосинтез новообразованной ДНК (стадию элонгации репликации ДНК), являются ДНК-полимеразы III. Имеются доказательства, что в димерной форме ДНК-полимераза III катализирует сопряженный синтез ведущей (лидирующей) и отстающей цепей ДНК при репликации.
ДНК-полимеразы I катализирует отщепление затравочного олигорибонуклеотидного праймера и заполнение образующихся после этого пробелов (ниш) дезоксирибонуклеотидами. ДНК-полимеразы II из Е. coli выполняет ≪ремонтные≫ функции, исправляя повреждения цепей ДНК.
Функцию раскручивания (расплетения) двойной спирали ДНК в репликационной вилке, происходящего за счет энергии гидролиза АТФ, выполняет специфический rep-белок, названный хеликазой. Образовавшиеся на определенное время одноцепочечные участки ДНК служат в качестве матрицы при репликации и стабилизируются при помощи особых белков, связывающихся с одноцепочечной ДНК (SSB-белки) и препятствующих обратному комплементарному взаимодействию цепей ДНК. В связи с этим их иногда называют дестабилизирующими двойную спираль белками. Имеются, кроме того, особые ферменты топоизомеразы (у прокариот одна из них названа ДНК-гиразой), которые играют особую роль в сверхспирализации, обеспечивая как репликацию, так и транскрипцию ДНК. Эти ферменты наделены способностью не только создавать супервитки, но и уничтожать суперспирализацию путем сшивания образующихся разрывов или разрезания ДНК.
В стадии инициации репликации ДНК участвует специфическая клеточная РНК-полимераза, названная праймазой, которая катализирует синтез короткого олигорибонуклеотида (от 10 до 60 нуклеотидов), т.е. праймера, с которого затем начинается синтез ДНК. В состав праймасомы входит также комплекс белков dna В и dna С, который вблизи репликационной вилки периодически участвует в формировании специфической вторичной структуры ДНК, подходящей для узнавания праймазой.
Важную функцию соединения двух цепей ДНК или замыкания двух концов одной цепи ДНК в процессе репликации либо репарации ДНК выполняют особые ферменты – ДНК-лигаза, катализирующая за счет энергии АТФ образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одной цепи и 5'-фосфатной группой другой цепи ДНК.
Билет №18
БИОСИНТЕЗ ХОЛЕСТЕРИНА
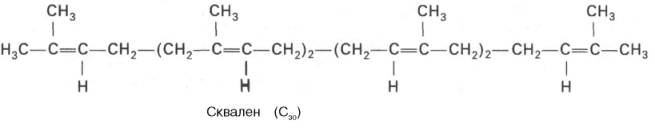
Сквален
Формулы ланостерина и холестерина
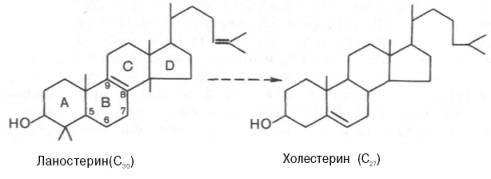
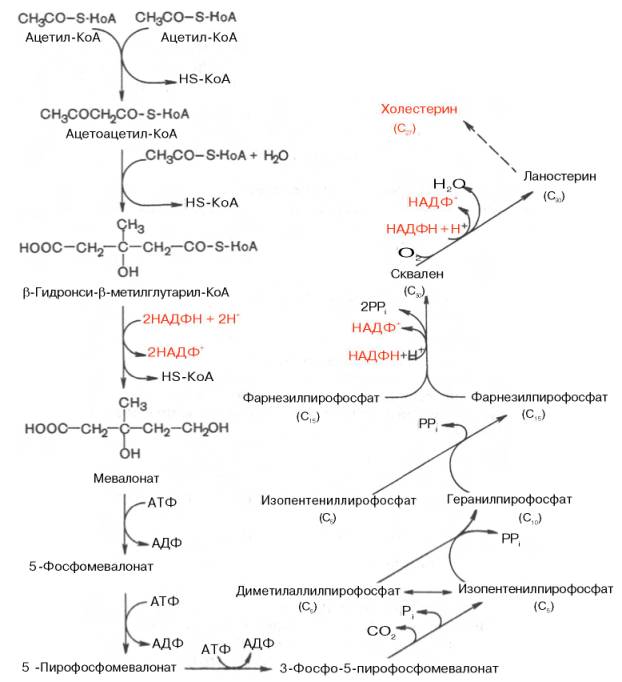
все атомы углерода холестерина происходят из ацетата. В синтезе холестерина можно выделить три основные стадии: I – превращение активного ацетата в мевалоновую кислоту, II – образование сквалена из мевалоновой кислоты, III – циклизация сквалена в холестерин.
ГМГ-КоА-редуктазная реакция – первая практически необратимая реакция в цепи биосинтеза холестерина. Она протекает со значительной потерей свободной энергии (около 33,6 кДж). Установлено, что данная реакция лимитирует скорость биосинтеза холестерина.
Начиная со сквалена, все промежуточные продукты биосинтеза холестерина (включая и холестерин) нерастворимы в водной среде. Поэтому они участвуют в конечных реакциях биосинтеза холестерина, будучи связанными со стеринпереносящими белками (СПБ). Это обеспечивает их растворимость в цитозоле клетки и протекание соответствующих реакций. Данный факт имеет важное значение и для вхождения холестерина в клеточные мембраны, окисления в желчные кислоты, превращения в стероидные гормоны. Как отмечалось, реакцией, регулирующей скорость биосинтеза холестерина в целом, является восстановление β-гидрокси-β-метилглутарил-КоА в мевалоновую кислоту, катализируемое ГМГ-КоА-редуктазой. Данный фермент испытывает регуляторное воздействие ряда факторов. В частности, скорость синтеза редуктазы в печени подвержена четким суточным колебаниям: максимум ее приходится на полночь, а минимум – на утренние часы. Активность ГМГ-редуктазы возрастает при введении инсулина и тире-оидных гормонов. Это приводит к усилению синтеза холестерина и повышению его уровня в крови. При голодании, тиреоидэктомии, введение глюкагона и глюкокорти-коидов, напротив, отмечается угнетение синтеза холестерина, что прежде всего связано со снижением активности ГМГ-КоА-редуктазы.