

2.Катаболизм триацилглицеролов. Реакции, механизм регуляции активности триглицеридлипазы. Нейрогуморальная регуляция липолиза (адреналин, глюкагон, инсулин).
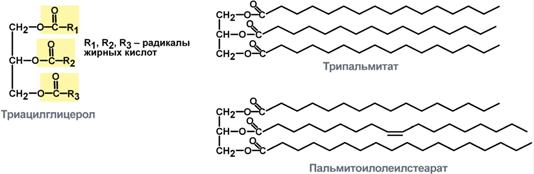
Триацилглицеролы (ТАГ, триглицериды, триацилглицерины, нейтральные жиры) являются наиболее распространенными липидами в организме человека. В состав ТАГ входит трехатомный спирт глицерол и три жирные кислоты. Жирные кислоты могут быть насыщенные (пальмитиновая, стеариновая) и мононенасыщенные (пальмитолеиновая, олеиновая). По строению можно выделить простые и сложные ТАГ. В простых ТАГ все жирные кислоты одинаковые, например трипальмитат, тристеарат. В сложных ТАГ жирные кислоты отличаются, например, дипальмитоилстеарат, пальмитоилолеилстеарат.
Путь катаболизма триацилглицеролов начинается с их гидролиза до жирных кислот и глицерола под действием липазы ; в основном этот процесс происходит в жировой ткани . Высвободившиеся жирные кислоты поступают в плазму крови , где связываются сывороточным альбумином . Затем свободные жирные кислоты переходят в ткани, где они либо окисляются, либо вновь подвергаются эстерификации . Ткани многих органов (печени, сердца, почек, мышц, легких, семенников, мозга), а также жировая ткань способны окислять длинноцепочечные жирные кислоты. Однако поступление этих кислот в клетки мозга затруднено. Что касается судьбы глицерола, то она зависит от того, присутствует ли в данной ткани необходимый активирующий фермент глицеролкиназа ( Биосинтез триацилглицеролов и фосфолипидов: метаболическая карта ). Значительное количество этого фермента обнаружено в печени , почках , кишечникче , бурой жировой ткани и в молочных железах в период лактации .
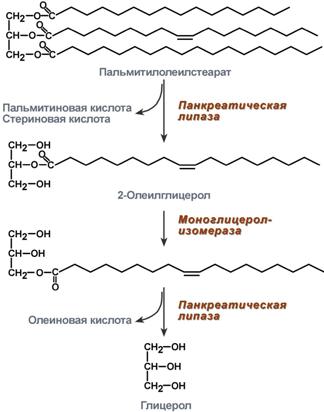
Переваривание ТАГ в кишечнике осуществляется под воздействием панкреатической липазы с оптимумом рН 8,0-9,0. В кишечник она поступает в виде пролипазы, активируемой при участии колипазы. Колипаза, в свою очередь, активируется трипсином и затем образует с липазой комплекс в соотношении 1:1. Панкреатическая липаза отщепляет жирные кислоты, связанные с С1 и С3 атомами углерода глицерола. В результате ее работы остается 2-моноацилглицерол (2-МАГ). 2-МАГ всасываются или превращаются моноглицерол-изомеразой в 1-МАГ. Последний гидролизуется до глицерола и жирной кислоты. Примерно 3/4 ТАГ после гидролиза остаются в форме 2-МАГ и только 1/4 часть ТАГ гидролизуется полностью.
В жировой ткани содержится несколько липаз, из которых наибольшее значение имеют триглицеридлипаза (так называемая гормоночувствитель-ная липаза), диглицеридлипаза и моноглицеридлипаза. Активность двух последних ферментов в 10–100 раз превышает активность первого. Три-глицеридлипаза активируется рядом гормонов (например, адреналином, норадреналином, глюкагоном и др.), тогда как диглицеридлипаза и мо-ноглицеридлипаза не чувствительны к их действию. Триглицеридлипаза является регуляторным ферментом.
Установлено, что гормоночувствительная липаза (триглицеридлипаза) находится в жировой ткани в неактивной форме, и активация ее гормонами протекает сложным каскадным путем, включающим участие по крайней мере двух ферментативных систем. Процесс начинается со взаимодействия гормона с клеточным рецептором, в результате чего модифицируется структура рецептора (сам гормон в клетку не поступает) и такой рецептор активирует аденилатциклазу (КФ 4.6.1.1). Последняя, как известно, катализирует образование циклического аденозинмонофосфата (цАМФ) из аденозинтрифосфата (АТФ):
![]()
Образовавшийся цАМФ активирует фермент протеинкиназу (КФ 2.7.1.37), который путем фосфорилирования неактивной триглицеридлипазы превращает ее в активную форму (рис. 11.1). Активная триглицеридлипаза расщепляет триглицерид на диглицерид и жирную кислоту. Затем при действии ди- и моноглицеридлипаз образуются конечные продукты липо-лиза – глицерин и свободные жирные кислоты, которые поступают в кровяное русло. Скорость липолиза триглицеридов не является постоянной, она подвержена регулирующему влиянию различных факторов, среди которых особое значение имеют нейрогормональные. Связанные с альбуминами плазмы крови в виде комплекса свободные жирные кислоты с током крови попадают в органы и ткани, где комплекс распадается, а жирные кислоты подвергаются либо β-окислению, либо частично используются для синтеза триглицеридов, глицерофосфолипидов, сфингофосфолипидов и других соединений, а также на эстерификацию холестерина.

Рис. 11.1. Липолитический каскад (по Стайнбергу).
ТГ - триглицериды; ДГ - диглицериды; МГ - моноглицериды; ГЛ - глицерин; ЖК - жирные кислоты.
Обмен липидов регулируется ЦНС. Кора большого мозга оказывает трофическое влияние на жировую ткань либо через нижележащие отделы ЦНС – симпатическую и парасимпатическую системы, либо через эндокринные железы. В настоящее время установлен ряд биохимических механизмов, лежащих в основе действия гормонов на липидный обмен.
Известно, что длительный отрицательный эмоциональный стресс, сопровождающийся увеличением выброса катехоламинов в кровяное русло, может вызвать заметное похудание. Уместно напомнить, что жировая ткань обильно иннервируется волокнами симпатической нервной системы, возбуждение этих волокон сопровождается выделением норадреналина непосредственно в жировую ткань. Адреналин и норадреналин увеличивают скорость липолиза в жировой ткани; в результате усиливается мобилизация жирных кислот из жировых депо и повышается содержание неэстерифи-цированных жирных кислот в плазме крови. Как отмечалось, тканевые липазы (триглицеридлипаза) существуют в двух взаимопревращающихся формах, одна из которых фосфорилирована и каталитически активна, а другая – нефосфорилирована и неактивна. Адреналин стимулирует через аденилатциклазу синтез цАМФ. В свою очередь цАМФ активирует соответствующую протеинкиназу, которая способствует фосфорилированию липазы, т.е. образованию ее активной формы. Следует заметить, что действие глюкагона на липолитическую систему сходно с действием кате-холаминов.
Не подлежит сомнению, что секрет передней доли гипофиза, в частности соматотропный гормон, оказывает влияние на липидный обмен. Гипофункция железы приводит к отложению жира в организме, наступает гипофизарное ожирение. Напротив, повышенная продукция СТГ стимулирует липолиз, и содержание жирных кислот в плазме крови увеличивается. Доказано, что стимуляция липолиза СТГ блокируется ингибиторами синтеза мРНК. Кроме того, известно, что действие СТГ на липолиз характеризуется наличием лаг-фазы продолжительностью около 1 ч, тогда как адреналин стимулирует липолиз почти мгновенно. Иными словами, можно считать, что первичное действие этих двух типов гормонов на липолиз проявляется различными путями. Адреналин стимулирует активность аденилатциклазы, а СТГ индуцирует синтез данного фермента. Конкретный механизм, с помощью которого СТГ избирательно увеличивает синтез аденилатциклазы, пока неизвестен.
Инсулин оказывает противоположное адреналину и глюкагону действие на липолиз и мобилизацию жирных кислот. Недавно было показано, что инсулин стимулирует фосфодиэстеразную активность в жировой ткани. Фосфодиэстераза играет важную роль в поддержании постоянного уровня цАМФ в тканях, поэтому увеличение содержания инсулина должно повышать активность фосфодиэстеразы, что в свою очередь приводит к уменьшению концентрации цАМФ в клетке, а следовательно, и к образованию активной формы липазы.