

Билет №26
1.Реплика́ция ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой
УОТСОНА—КРИКА МОДЕЛЬ ДНК, согласно к-рой молекула ДНК состоит из двух антипараллельных полинуклеотидных цепей, образующих правильную правозакрученную перевитую спираль и удерживаемых имеете водородными связями за счёт взаимодействия пар азотистых оснований.
В процессе репликации двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных цепей, раскручивается на отдельные цепи и одновременно начинается синтез новых полинуклеотидных цепей; при этом исходные цепи ДНК играют роль матриц. Новая цепь, синтезирующаяся на каждой из исходных цепей, идентична др. исходной цепи. Когда процесс завершается, образуются две идентичные двойные спирали, каждая из к-рых состоит из одной старой (исходной) и одной новой цепи. Таким образом от одного поколения к другому передается только одна из двух цепей, составляющих исходную молекулу ДНК,-т. наз. полуконсервативный механизм репликации.
Репликация состоит из большого числа последоват. этапов, к-рые включают узнавание точки началу репликации, расплетание исходного дуплекса (спирали), удержание его цепей в изолированном друг от друга состоянии, инициацию синтеза на них новых дочерних цепей, их рост (элонгацию), закручивание цепей в спираль и терминацию (окончание) синтеза. Все эти этапы репликации, обеспечивает ДНК-репликазная система, или реплисома. Функцион. единица репликации-репликон, представляющий собой сегмент (участок) хромосомы или внехромосомной ДНК, ограниченный точкой начала, в к-рой инициируется репликация, и точкой окончания, в к-рой репликация останавливается. Скорость репликации контролируется на стадии инициации. Эукариотич. хромосомы содержат большое число репликонов, каждый из к-рых также однократно инициируется за один клеточный цикл.
Начиная с точки инициации, репликация осуществляется в ограни-ченной зоне, перемещающейся вдоль исходной спирали ДНК. Эта активная зона репликации (т. наз. репликац. вилка) может двигаться в обоих направлениях. В ходе репликации рост цепи осуществляется благодаря взаимод. дезоксирибонуклеозидтрифосфата с 3'-ОН концевым ну-клеотидом уже построенной части ДНК; при этом отщепляется пирофосфат и образуется фосфодиэфирная связь. Рост полинуклеотидной цепи идет только с ее З'-конца, т. е. в направлении 5' : 3' . Фермент, катализирующий эту р-цию,-ДНК-полиме-раза -не способен начать матричный синтез на одноцепочечной ДНК, если нет хотя бы олигонуклеотидного биспирального участка (т. наз. затравочного олигонуклеотида) комплементарного матрице; затравочным олигонуклеотидом во мн. случаях является не ДНК, а РНК. Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в цепи ДНК, обеспечивается расщеплением фосфатной связи между - и -фосфатными группами нуклеозидтрифосфата.
ДНК-полимераза имеет один центр связывания нуклеозидтрифосфата, общий для всех четырех нуклеотидов. Выбор из среды нуклеотида, основание к-рого комплементарно очередному основанию матрицы, протекает без ошибок, благодаря определяющему влиянию ДНК-матрицы (исходной цепи ДНК). Репликац. вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится непрерывно, а другая-с перерывами. Первую наз. ведущей, или лидирующей, цепью, а вторую-отстающей. Синтез второй цепи идет медленнее; хотя в целом эта цепь строится в направлении 3' : 5', каждый из ее фрагментов в отдельности наращивается в направлении 5' : 3'. Благодаря такому прерывистому механизму синтеза, репликация обеих антипараллельных цепей осуществляется с участием одного фермента-ДНК-полимеразы, катализирующего наращивание нуклеотидной цепи только в направлении 5' : 3'.
В качестве затравок для синтеза фрагментов отстающей цепи служат короткие отрезки РНК, комплементарные матричной цепи ДНК. Эти РНК-затравки (праймеры), синтезируются на матрице отстающей цепи из рибонуклеозидтрифосфатов в направлении 5' : 3' с помощью фермента РНК-праймазы. РНК-праймеры затем наращиваются дезоксинуклеотидами с 3'-конца ДНК-поли-меразой, к-рая продолжает наращивание до тех пор, пока строящаяся цепь не достигает РНК-затравки, присоединенной к 5'-концу предыдущего фрагмента. Образующиеся таким образом фрагменты (т. наз. фрагменты Оказаки). Чтобы обеспечить образование непрерывной цепи ДНК из многих таких фрагментов, в действие вступает особая система репарации ДНК, удаляющая РНК-затравку и заменяющая ее на ДНК. Завершает весь процесс фермент ДНК-лигаза, катализирующий образование фосфодиэфирной связи между группой З'-ОН нового фрагмента ДНК и 5'-фосфатной группой предыдущего фрагмента. Образование этой связи требует затраты энергии.
Раскручивание двойной спирали и пространств. разделение цепей осуществляется белками. Геликазы расплетают короткие участки ДНК, находящиеся непосредственно перед репликац. вилкой. На разделение каждой пары оснований расходуется энергия гидролиза двух молекул АТФ до аденозиндифосфата и фосфата. К каждой из разделившихся цепей присоединяется неск. молекул ДНК-связывающих белков, к-рые препятствуют образованию комплементарных пар и обратному воссоединению цепей. Благодаря этому нуклеотидные последовательности цепей ДНК оказываются доступными для репликативной системы. Др. специфич. белки помогают праймазе получить доступ к матрице отстающей цепи. В результате праймаза связывается с ДНК и синтезирует РНК-затравки для фрагментов отстающей цепи. Для формирования новых спира-,лей не требуется ни затрат энергии, ни участия к.-л. "закручивающего" фермента.
2. Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитами являются в первую очередь молочная и пи-ровиноградная кислоты, так называемые гликогенные аминокислоты, гли-церол и ряд других соединений. Иными словами, предшественниками глюкозы в глюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот. У позвоночных наиболее интенсивно глюконеогенез протекает в клетках печени и почек (в корковом веществе). Большинство стадий глюконеогенеза представляет собой обращение реакции гликолиза. Только 3 реакции гликолиза (гексокиназная, фосфо-фруктокиназная и пируваткиназная) необратимы, поэтому в процесс глю-конеогенеза на 3 этапах используются другие ферменты. Рассмотрим путь синтеза глюкозы из пирувата.
Образование фосфоенолпирувата из пирувата. Синтез фосфоенолпирувата осуществляется в несколько этапов. Первоначально пируват под влияние пируваткарбоксилазы и при участии СО2 и АТФ карбоксилируется с образованием оксалоацетата:
1
Первый этап синтеза протекает в митохондриях. Пируват-карбоксилаза, которая катализирует эту реакцию, является аллостери-ческим митохондриальным ферментом. В качестве аллостерического активатора данного фермента необходим ацетил-КоА. Мембрана митохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же, в митохондриях, восстанавливается в малат:
2
Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. В митохондриях отношение НАДН/НАД+ относительно велико, в связи с чем внутримитохондриальный оксалоацетат легко восстанавливается в малат, который легко выходит из митохондрии через митохондриальную мембрану. В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется при участии цитоплазматической НАД-за-висимой малатдегидрогеназы:
3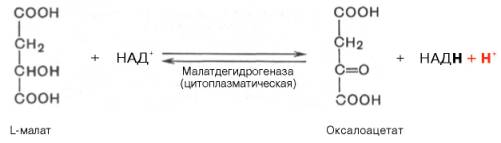
Затем оксалоацетат в результате декарбоксилирования и фосфорили-рования под влиянием фермента фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват. Донором фосфатного остатка в реакции служит гуанозинтрифосфат (ГТФ):
4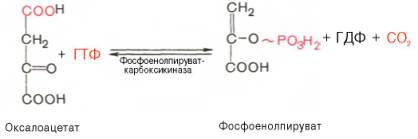
Далее идут обратимые реакции гликолиза:
1
2
3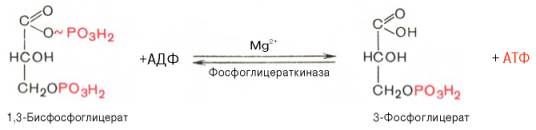
4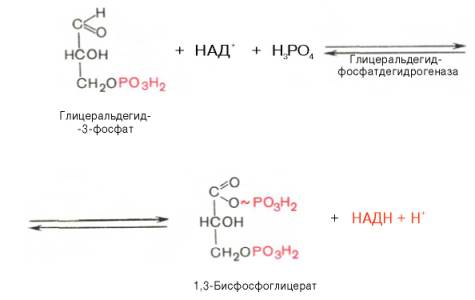
5
6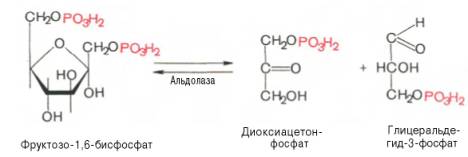
7. Далее следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в обход этой эндергонической реакции:
![]()
8.
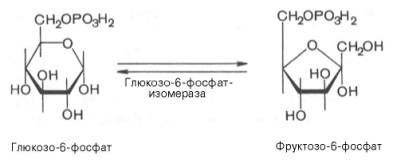
9. Образование глюкозы из глюкозо-6-фосфата.
![]()