

Билет 19
1.БИОСИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ И ФОСФОГЛИЦЕРИДОВ. Биосинтез триацилглицеролов и глицеролфосфатидов начинается с общих предшественников. В животных тканях биосинтез триацилглицеролов и главных фосфолипидов - фосфатидилэтаноламина и фосфатидилхолина начинается с двух общих предшественников и имеет несколько общих этапов. Общими предшественниками служат СоА-эфиры жирных кислот и глицерол-3-фосфат. Глицеролфосфат может образовываться двумя путями. В ходе гликолиза он возникает из дигидроксиацетонфосфата под действием цитоплазматического NAD-зависимого фермента глицеролфосфатдегидрогеназы: Дигидроксиацетонфосфат + NADH + Н+ ↔ L-глицерол-З-фосфат + NAD + .
Кроме того, глицеролфосфат может образоваться из глицерола под действием глицеролкиназы: АТР + Глицерол → Глицерол-3-фосфат + ADP. Другими предшественниками триацил-глицеролов служат СоА-производные жирных кислот, образующиеся при помощи ацил-СоА—синтетаз. Жирная кислота + АТР + СоА—SH — Ацил—S—СоА + AMP + РРi На первом этапе биосинтеза триацил-глицеролов происходит ацилирование двух свободных гидроксильных групп глицеролфосфата двумя молекулами Со-А-производных жирных кислот с образованием диацилглицерол-3-фосфата: Ацил—S—СоА + Глицерол → Моноацилглицерол-3-фосфат + СоА—SH Моноацилглицерол-3-фосфат + + Ацил—S—СоА → Диацилглицерол-3-фосфат + СоА—SH
Д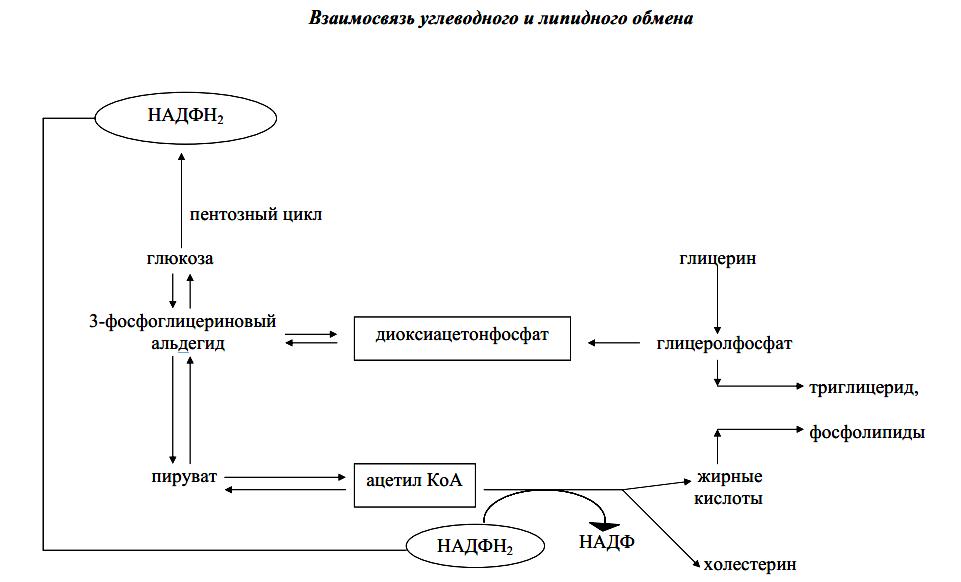 иацилглицерол-3-фосфат
(чаще его называют фосфатидной
кислотой) встречается в клетках
лишь в следовых количествах, однако он
является важным промежуточным продуктом
в биосинтезе липидов. В ходе синтеза
триацилглицеролов фосфатидат
гидролизуется фосфатидат-фосфатазой с
образованием 1,2-диа-цилглицерола:
Фосфатидат
+ Н20 → 1,2-диацилглицерол + Рi.
Диацилглицерол,
взаимодействуя с третьей молекулой
СоА-производного жирной кислоты,
превращается затем в
триацилглицерол:
СоА-производное
жирной кислоты +1,2-диацилглицерол →
Триацилглиперол + СоА—SH.
Формирование
каждой эфирной связи триацилглицеролов
требует значительного количества
свободной энергии. Для того чтобы
возникла эфирная связь, жирная кислота
сначала должна активироваться путем
образования эфира с СоА; для этой реакции
необходима энергия двух высокоэнергетических
фосфатных связей, так как она протекает
благодаря пирофосфатному расщеплению
АТР и последующему гидролизу образовавшегося
пирофосфата.
иацилглицерол-3-фосфат
(чаще его называют фосфатидной
кислотой) встречается в клетках
лишь в следовых количествах, однако он
является важным промежуточным продуктом
в биосинтезе липидов. В ходе синтеза
триацилглицеролов фосфатидат
гидролизуется фосфатидат-фосфатазой с
образованием 1,2-диа-цилглицерола:
Фосфатидат
+ Н20 → 1,2-диацилглицерол + Рi.
Диацилглицерол,
взаимодействуя с третьей молекулой
СоА-производного жирной кислоты,
превращается затем в
триацилглицерол:
СоА-производное
жирной кислоты +1,2-диацилглицерол →
Триацилглиперол + СоА—SH.
Формирование
каждой эфирной связи триацилглицеролов
требует значительного количества
свободной энергии. Для того чтобы
возникла эфирная связь, жирная кислота
сначала должна активироваться путем
образования эфира с СоА; для этой реакции
необходима энергия двух высокоэнергетических
фосфатных связей, так как она протекает
благодаря пирофосфатному расщеплению
АТР и последующему гидролизу образовавшегося
пирофосфата.
2.Биосинтез пуриновых нуклеотидов
Первая специфическая реакция образования пуриновых нуклеотидов - перенос амидной группы Глн на ФРДФ с образованием 5-фосфорибозил-1 -амина. Эту реакцию катализирует фермент амидофосфорибозилтрансфераза. При этом формируется β-N-гликозидная связь. Затем к аминогруппе 5-фосфорибозил-1-амина присоединяются остаток глицина, N5,N10-метенил-Н4-фолата ещё одна амидная группа глутамина, диоксид углерода, аминогруппа аспартата и формильный остаток N10-формил Н4-фолата. Результатом этой десятистадийной серии реакций является образование первого пуринового нуклеотида - инозин-5'-монофосфата (ИМФ), на синтез которого затрачивается не менее шести молекул АТФ.
ИМФ(иозиновая кислота) в основном используется на синтез АМФ или ГМФ. Небольшое количество этого продукта обнаруживается также в тРНК в качестве одного из минорных нуклеотидов.
Превращение ИМФ в АМФ и ГМФ в обоих случаях включает 2 стадии и идёт с затратой энергии. Аденилосукцинатсинтетаза, используя энергию ГТФ, присоединяет аспартат к ИМФ с образованием аденилосукцината, который в реакции, катализируемой аденилосукциназой, отщепляет фумарат и превращается в АМФ.
Второй пуриновый нуклеотид (ГМФ) образуется также в 2 стадии. Сначала ИМФ окисляется NAD+-зависимой ИМФ-дегидрогеназой с образованием ксантозин-5'-монофосфата (КМФ). Последующее трансамидирование гидроксильной группы при С2-пуринового кольца КМФ катализирует ГМФ-синтетаза с использованием амидной группы Глн и энергии АТФ.
При образовании пуриновых нуклеотидов ГТФ расходуется на синтез АМФ, а АТФ - на синтез ГМФ. Перекрёстное использование пуриновых нуклеозидтрифосфатов на образование конечных продуктов синтеза помогает поддерживать в клетках баланс адениловых и гуаниловых нуклеотидов.
Огромные затраты энергии для синтеза пуриновых нуклеотидов de novo не способны полностью обеспечить субстратами синтез нуклеиновых кислот в период гаструляции и раннего роста ребёнка. Потребность в большом количестве нуклеотидов привела к развитию "запасных" путей синтеза этих молекул. Наибольшее значение в этом процессе имеют ферменты, осуществляющие превращение пуринов в мононуклеотиды с использованием ФРДФ как донора остатка фосфорибозы.
Основным показателем, от которого зависит синтез пуриновых нуклеотидов, служит концентрация ФРДФ, которая, в свою очередь, зависит от скорости его синтеза, утилизации и разрушения. Количество ФРДФ определяется доступностью рибозо-5-фосфата и активностью ФРДФ синтетазы - фермента, чувствительного к концентрации фосфата и пуриновых нуклеотидов. Внутриклеточная концентрация ФРДФ строго регулируется и обычно низкая. ФРДФ синтетаза - аллостерический фермент. Он активируется неорганическим фосфатом (Рi) и ингибируется пуриновыми нуклеозид- моно-, ди- и трифосфатами. ФРДФ служит не только субстратом, но и аллостерическим активатором второй реакции синтеза пуринонуклеотидов de novo, которую катализирует амидофосфорибозилтрансфераза.
Пуриновые нуклеотиды, особенно АМФ и ГМФ по механизму отрицательной обратной связи ингибируют амидофосфорибозилтрансферазу, которая катализирует первую специфическую реакцию синтеза пуриновых нуклеотидов de novo.
Метаболическая цепь образования АМФ и ГМФ de novo регулируется также в месте её разветвления: АМФ ингибирует аденилосукцинатсинтетазу, а ГМФ - реакцию образования ксантиловой кислоты, которую катализирует ИМФ дегидрогеназа. Перекрёстная регуляция путей использования ИМФ служит для того, чтобы снизить синтез одного пуринового нуклеотида при дефиците другого.
Помимо ферментов основного пути синтеза пуриновых нуклеотидов de novo, регулируется также активность ферментов "запасных" путей: аденинфосфорибозилтрансфераза ингибируется АМФ, а гипоксантин-гуанинфосфорибозилтрансфераза - ИМФ и ГМФ.
