

(ионизаций, возбуждений, упругих и неупругих атомных столкновений) в образовании первичных повреждений в клетках, по крайней мере, с точки зрения образования структурных дефектов. Однако полная картина физического действия излучения на биологические объекты в каждом конкретном случае должна включать в себя, как отмечалось выше, не только все первичные повреждения, но и информацию об их распределении в объеме.
Исходными для весьма простого количественного описания распределения дефектов по пространству могут быть полученные ранее выражения для числа структурных дефектов (7.15) и (7.22). Непосредственно из формулы (7.15) вытекает, например, выражение для пространственного распределения структурных дефектов в сердцевине трека при прохождении через биологический объект быстрого тяжелого иона
dN (i) δ(r ) |
m1 |
n2 (z1 z2e2 )2 |
1 |
1 |
|
|
δ(r ) |
|
|||
δ(r ) dx 2πr |
=π m |
|
|
|
|
|
|
|
|
2πr . |
(8.15) |
E |
(x) |
|
−α E |
|
|||||||
T |
(x) |
||||||||||
|
2 |
1 |
|
|
d |
1 1 |
|
|
|
|
|
Здесь в явном виде учтена зависимость текущей энергии иона от длины пути, а распределение в поперечном к треку направлении выбрано в виде дельтафункции (бесконечно тонкий шнур). Зависимость энергии иона от координаты можно получить, используя известную связь между длиной пробега и энергией RL (E0 ) , которая была получена ранее. Для этого нужно в выражение
(5.18) в качестве нижнего предела подставить величину E1 .
Аналогичным образом из (7.22) можно получить и поперечное к оси трека распределение структурных дефектов. Однако в самом простом приближении для этого достаточно считать, что радиус сердцевины определяется длиной пробега δ -электронов с энергией 100-400эВ, а радиус, например, δ -электронного облака – длиной пробега δ -электронов с
максимальной энергией Tm ≈ 2mev12 . Аналогичным образом, максимальный
радиус области образования структурных дефектов в окрестности трека связан с длиной пробега протонов отдачи и первичных выбитых атомов с максимальной полученной от иона энергией. Понятно, что с ростом скорости полное число вторичных частиц, выбиваемых на единице пути, убывает
обратно пропорционально v12 , поэтому пространственная плотность числа
вторичных частиц очень быстро уменьшается с ростом скорости по мере увеличения радиуса облака и уменьшения числа частиц в нем.
7.4Принципы попадания и мишени в радиобиологии
Всоответствии с изложенной в разделе 1 иерархией уровней радиационных и радиобиологических эффектов, естественной является классификация радиобиологических моделей по уровням биологической организации: химическому, биохимическому и клеточному, что составляет предмет радиобиологии. Далее будут кратко рассмотрены только так
называемые физические модели, которые исторически были предложены раньше других и являются развитием теории мишени. Физическими эти модели названы потому, что они непосредственно связывают первичные физические процессы с конечными биологическими эффектами, минуя промежуточные стадии. Естественно, что такой подход при моделировании, во-первых, существенно ограничивает применимость физических моделей и, во-вторых, не дает динамической картины процессов на других уровнях.
Физические модели можно разделить на четыре группы: теория мишени и ее модификации, двухкомпонентные модели, теория дуального действия и ее модификации и некоторые другие вероятностные модели. Среди этих моделей, в принципе, есть такие, которые рассматривают эффекты на двух или более уровнях организации биологического объекта. Например, теория дуального действия может быть отнесена к более сложным системным моделям, однако она не учитывает динамику внутриклеточных процессов, не идентифицирует однозначно первичные повреждения с какими-либо известными повреждениями (например, одноили двунитевыми разрывами ДНК) и непосредственно связывает первичные процессы передачи энергии излучения с конечными повреждениями (аберрация хромосом). По этим признакам теория дуального действия относится к физическим моделям.
В основу всех физических моделей заложены фундаментальные принципы попадания и мишени. Первый принцип утверждает, что энергия падающих частиц передается веществу в пространстве и во времени статистически независимыми дискретными порциями, вследствие чего в очень малых микрообъемах реально выделенная удельная энергия может во много раз отличаться от средней поглощенной энергии в объеме. Принцип мишени отражает, во-первых, факт крайней неоднородности (с физической, химической и биологической точек зрения) живых объектов (организмов, тканей, клеток) и, во-вторых, различную значимость разных микрообъемов и структур для функционирования клетки и, в более широком плане, всего биологического объекта.
Принципы попадания и мишени имеют всеобщее значение и по существу должны применяться всегда ввиду стохастичности процессов передачи энергии биологическим объектам и различной значимости для их существования и функционирования характерных неоднородностей. В самом общем случае эти принципы формулируются в следующих предположениях.
1. Биологический объект содержит m чувствительных областей (мишеней), причем при n или более событий попадания мишень с большой вероятностью инактивируется (гибнет или теряет определенные свойства и способности, которые могут быть измерены количественно). Если число событий попадания меньше n , мишень остается без изменений (принцип попадания).
2. События, которые произошли в объекте, но вне мишеней, не изменяют свойств мишеней и соответственно свойств всего биологического
объекта (принцип мишени).
3.Под действием излучения биологический объект в целом инактивируется с большой вероятностью, если поражаются k или более из m мишеней (свойство объекта).
4.События передачи биологическому объекту энергии (попадания) статистически распределены в пространстве и во времени, не зависят друг от друга и могут быть описаны в рамках физических моделей взаимодействия излучения с веществом (принцип попадания).
Следует заметить, что вводимые таким образом мишени вообще говоря не представляют собой непосредственно какие-либо чувствительные к облучению структуры объекта либо его элементы. По существу здесь имеется
ввиду некоторый эффективный объем, поглощенная энергия из которого может достигнуть чувствительной структуры и вызвать первичное поражение последней. Поэтому по объему мишени могут быть как больше, так и меньше объемов конкретных структур. Размеры мишени зависят от многих причин (качества излучения, состояния объекта и т.д.).
Далее, в физической модели попадания и мишени постулируется статистическая вероятность выживания одной мишени в виде
S |
|
n−1 |
(Vn )p |
(8.16) |
= exp(−Vn )∑ |
r . |
|||
1 |
r |
p=0 |
p! |
|
|
|
|
||
Здесь экспоненциальный характер выживания выбран на основании множества экспериментальных данных. В соответствии с выражением (8.16) при n → ∞ выживаемость одной мишени S1 →1, что и понятно, поскольку
невозможно инактивировать мишень, для поражения которой требуется бесконечное число попаданий. С уменьшением параметра n выживаемость мишени во все большей мере определяется экспоненциальной функцией, в которой величина Vnr – это среднее число попаданий падающих частиц в
мишень, V – радиочувствительный объем мишени и nr - число попаданий на
единицу объема биологического объекта. Заметим, что при очень большом среднем числе попаданий Vnr >>1 в мишень вероятность ее выживания мала,
однако не равна нулю при любом n .
Если для инактивации всего объекта требуется поражение любых k мишеней из m возможных, то статистически вероятность его выживания описывается выражением
k−1 |
m! |
S m−l (1 |
− S )l . |
|
|
S = ∑ |
(8.17) |
||||
|
|||||
l=0 |
(m −l)!l! 1 |
1 |
|
||
Не приводя соответствующего вывода формулы для кривой выживаемости S , которая сводится к числу сочетаний из m элементов множества по l , рассмотрим два предельных случая, которые показывают ее правильность. Если для инактивации объекта достаточно поразить хотя бы одну мишень из
m , то при k =1 из (8.17) получаем
S = S m . |
(8.18) |
1 |
|
Таким образом, в этом случае вероятность выживания объекта в целом равна произведению вероятностей выживания каждой из мишеней. В обратном предельном случае, когда необходимо поражение всех m мишеней внутри объекта, из (8.17) после подстановки k = m получаем
S = ∑ m! |
S m−l (1− S )l − (1− S )m =1− (1− S )m . |
(8.19) |
||||
m |
|
|
|
|
|
|
|
1 |
1 |
1 |
|
||
l=0 |
(m −l)!l! |
1 |
|
|||
Отсюда следует, что вероятность выживания объекта отличается от единицы на величину, равную вероятности одновременного поражения всех мишеней.
Если для инактивации объекта необходимо поражение всех мишеней, а для инактивации мишени достаточно только одного события попадания, то, подставляя выражение (8.16) при n =1 в (8.19), получаем еще одну частную формулу, вытекающую из общей формулы (8.17):
S =1−[1−exp(−Vn )]m . |
(8.20) |
r |
|
Это соотношение оказалось весьма работоспособным и его часто используют в радиобиологии для описания простых случаев радиационного поражения. Наконец, если в объекте существует всего одна мишень, для инактивации которой достаточно одного попадания, то непосредственно из (8.20) при m =1 получаем
S = exp(−Vnr ) . |
(8.21) |
Тогда кривая выживаемости S , показывающая долю выживших по выбранному признаку биологических объектов, представляет собой обычную экспоненту.
В более сложных модификациях теории мишени можно учесть случаи, при которых для одной и той же группы объектов (популяции) меняются от одного объекта к другому число мишеней, число попаданий или объем мишеней. Кроме того, для реальных мишеней более правдоподобно, что поражение мишеней происходит при различных значениях n (и даже любых n ), но с разной вероятностью, зависящей от числа n . В действительности эти случаи достаточно хорошо описываются даже в рамках представленной теории. Например, если может варьировать от мишени к мишени число n , то под n в выражении (8.16) можно понимать некоторое среднее значение числа необходимых попаданий. Это тем более оправдано, что, как отмечалось выше, при любом даже наперед заданном n статистическая вероятность выживания мишени всегда отлична от нуля и может быть описана выражением (8.16). В целом представленная теория попаданий и мишени в ее самом общем представлении (8.17) способна объяснить очень большое число самых разнообразных кривых выживаемости. Иное дело, что при большом
количестве параметров модели практическая ценность ее применения понижается из-за большей неоднозначности, возникающей при интерпретации экспериментальных данных. Принципиальный же недостаток теории попадания и мишени заключается в привязывании биологического эффекта непосредственно к физическому уровню и характерным для него актам первичного физического действия.
Рассмотрим применение теории мишени для описания некоторых радиобиологических эффектов. Наиболее плодотворные результаты были получены с помощью простых вариантов этой теории при описании инактивации малых биологических объектов, типа ферментов, генов, вирусов и фагов. Исходя из экспериментальных данных, согласно которым вероятность выживания этих объектов в зависимости от дозы описывается простой экспонентой, можно предположить, что для инактивации таких объектов достаточно одного события попадания в единственный радиочувствительный объем внутри объекта. В соответствии с этим перейдем к описанию с помощью дозы излучения и запишем для самого простого варианта теории, представленного выражением (8.21), вероятность выживания в виде
S = exp(−αD) , |
(8.22) |
где α - чувствительность объекта на единицу дозы D , так что величина α−1 = D0 – это такая полная доза облучения, при которой выживаемость
уменьшается в e раз.
С другой стороны, в соответствии с выражением (1.27) величину Vnr можно выразить через дозу облучения Dr , поглощаемую непосредственно в той реакции, которая собственно и является событием попадания в мишень. В
результате получаем |
Dr ρV |
|
|
|
n V = |
, |
(8.23) |
||
|
||||
r |
T r |
|
|
|
|
|
|
где  T
T  r - средняя передача энергии падающей частицы при попадании в
r - средняя передача энергии падающей частицы при попадании в
радиочувствительный объем мишени. В данном случае использование выражений типа (1.27) или (8.11) является корректным потому, что речь идет о заведомо малых радиочувствительных объемах, которые можно описывать аналогично тонким слоям вещества. Кроме того, здесь для простоты не учитываются дополнительные возможные каналы реакции. Например такие, которые связаны с первичными выбитыми атомами и δ -электронами.
Понятно, что доза Dr , как и соответствующая часть тормозной
способности частицы, представляет собой только часть полной дозы D излучения, поэтому выражение (8.23) можно переписать в виде
n V = |
ρV |
δ |
D , |
(8.24) |
|
|
|||||
r |
T |
|
r |
|
|
|
r |
|
|
||
где δr - доля потерь энергии падающей частицы, связанных с конкретным физическим механизмом создания структурного дефекта, а величина  T
T 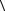 r
r
представляет собой среднюю передачу энергии в таком процессе попадания. Благодаря выражению (8.24) можно в явном виде записать чувствительность объекта на единицу дозы:
α = D −1 |
= |
ρV |
δ |
. |
(8.25) |
|
|
||||||
0 |
|
T |
|
r |
|
|
|
|
r |
|
|
||
Яснее увидеть физический смысл выражения (8.25) можно, если выбрать какой-либо конкретный механизм попадания. Пусть событие попадания – это образование структурного дефекта в молекулах определенного типа объекта, например, разрыв в цепочках ДНК. Чтобы
получить численные оценки, заменим величину 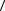 T
T 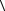 r −1 ее значением (7.19) для тяжелых ионов, тогда выражение (8.25) приобретает вид
r −1 ее значением (7.19) для тяжелых ионов, тогда выражение (8.25) приобретает вид
−1 |
|
ρV α1E1 −Td |
|
|
|
|||||
α = D0 |
≈ |
|
|
|
δЯ (E1 ) . |
(8.26) |
||||
T |
α E ln(α E T ) |
|||||||||
|
|
d |
1 1 |
|
1 |
1 |
d |
|
|
|
Теперь можно ввести величину |
|
T |
0 |
=δ |
−1 |
(E )T , которая |
характеризует |
|||
|
|
|
|
|
|
Я |
1 |
d |
|
|
потерю энергии частицы с учетом всех физических механизмов взаимодействия, включая неупругие, при заданной передаче энергии в одном ядерном столкновении. В результате получаем окончательное выражение для чувствительности на единицу дозы
|
−1 |
|
|
ρV |
|
α1E1 −Td |
|
|
|
|||
α |
= D0 |
|
≈ |
|
|
|
|
|
|
|
. |
(8.27) |
|
|
T 0 |
α1E1 ln(α1E1 |
|
Td ) |
|||||||
|
|
|
|
|
|
|
|
|||||
Легко видеть, что в предельном случае очень больших скоростей |
||||||||||||
движения и энергий E |
>>α |
−1T |
величина δ |
Я |
(E ) уменьшается |
с ростом |
||||||
1 |
1 |
|
d |
|
|
|
|
1 |
|
|
||
неупругих потерь, а передача энергии в одном столкновении возрастает. Иными словами, возрастают неупругие потери энергии тяжелого иона в условиях, когда число ядерных столкновений уменьшается, а передача энергии в каждом из них возрастает и становится много больше минимально нужной для создания дефекта энергии Td . Если полностью пренебречь
возможностью образования каскадных процессов, то оба этих фактора приводят к уменьшению чувствительности α и возрастанию дозы излучения, необходимой для уменьшения выживаемости в e раз.
Наоборот, при малых скоростях и энергиях налетающего иона, когда
E →α |
−1T |
и возрастает доля ядерных потерь δ |
Я |
(E ) ~1, величина |
T |
0 |
|
1 |
1 |
d |
|
1 |
|
||
уменьшается, поскольку увеличивается число ядерных столкновений на единице пути, а передаваемая в них энергия приближается к Td . В результате
при малых скоростях, когда тормозная способность – электронная и ядерная,

резко возрастают, величина α |
стремится |
|
к |
конечному |
пределу, |
|||||
определяемому эффективной энергией |
T |
0 |
=δ |
−1 (E )T |
~T . |
|
|
|||
|
|
|
|
Я |
1 |
d |
d |
|
|
|
Последнее |
обстоятельство |
показывает, |
|
|
почему |
с |
учетом |
|||
экспериментальных данных говорят о так называемом насыщении чувствительности при большой тормозной способности. Действительно,
средняя длина треков l в пределах объекта типа фага совпадает с размерами самого фага и составляет величину порядка 10-2мкм. При большой тормозной способности порядка 100кэВ/мкм это означает, что в пределах фага выделяется энергия около 1кэВ, которая во много раз превосходит энергию
порядка 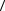 T
T  0 ~60эВ, необходимую для поражения фага. Таким образом, если
0 ~60эВ, необходимую для поражения фага. Таким образом, если
выразить чувствительность непосредственно через тормозную способность, то необходимо дополнительно вводить так называемый фактор перекрывания. При заданной поглощенной дозе фактор перекрывания призван учитывать меньшую в определенном смысле (из-за избытка выделяемой в объеме энергии) эффективность излучений с высокими значениями тормозной способности. В то же время, использование выражений (8.25) – (8.27), которые связывают чувствительность с энергией частицы, средней передачей энергии в одном событии попадания и долей передачи энергии в таких столкновениях, необходимости отдельно учитывать фактор перекрывания просто не возникает.
Итак, из выражения (8.27) следует, что вероятность выживания описывается в данном случае экспоненциальной зависимостью, в которой чувствительность с ростом тормозной способности стремится к некоторому
конечному |
предельному значению, которое связано с величиной порядка |
||||
T |
0 |
~60эВ. |
Зная объем объекта, можно рассчитать величину |
D =α−1 |
. При |
|
|
значения D0 , вычисленного в соответствии с |
0 |
|
|
сравнении |
(8.25), |
и из |
|||
экспериментальных кривых инактивации фагов, было получено хорошее согласие между ними для мелких фагов. Так, для фага S13, диаметр которого
равен |
1,6 10-2мкм, |
теоретическое |
значение |
D =4,67 103Гр, |
а |
|
|
|
|
0 |
|
экспериментальное находится в диапазоне 3,9 103 – 5,8 103Гр. Наоборот, зная D0 из эксперимента, можно определить чувствительный объем фага или
мелкого вируса. Как и следовало ожидать, из этих расчетов следует, что чувствительный объем в фаге приблизительно равен объему самого фага.
Причиной инактивации вирусов и фагов является, по-видимому, однонитевой разрыв или повреждение основания ДНК, для образования которых достаточно одного акта соответствующего первичного взаимодействия излучения с молекулой ДНК. Сечение инактивации даже при высоких S может быть много меньше сечения объекта, например, для больших вирусов. Это значит, что не весь объем объекта является в этом случае чувствительным. Для больших вирусов таким чувствительным объектом является ДНК, которая заключена внутри белковой