

Биосинтез пурииовых нуклеотидов
Методом меченых атомов в 30-е годы двадцатого века доказано, что пуриновые основания, образующиеся в процессе превращения, нуклеиновых кислот в кишечнике, в дальнейшем практически не используются. Поэтому пуриновая структура образуется из мелких фрагментов, поставляемых из низкомолекулярных соединений. Можно выделить следующие стадии синтеза пуриновых нуклеотидов:
1-я стадия - синтез β-5-фосфорибозил-1-амина на основе продукта пентозофосфатного цикла α-рибозо-5-фосфата:

-
α-рибозо-5-фосфат
α-рибозо-5-фосфорибозил-1-дифосфат (ФРДФ)
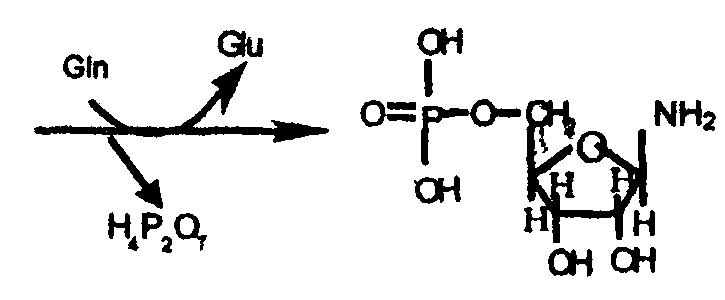
β-5-фосфорибозил-1-амин
2-я стадия - серия реакций, приводящая к формированию пуринового цикла и образованию инозиновой кислоты (ИМФ), пуриновая часть которой представлена гипоксантином:

Инозинмонофосфат (ИМФ, инозиновая кислота)
3-я стадия - синтез на основе ИМФ пуриновых нуклеотидов - АМФ и ГМФ, которые затем под действием соответствующих ферментов могут превращаться в нуклеозидди- и три фосфаты:

Биосинтез пиримидиновых нуклеотидов
Пиримидиновое ядро пиримидиновых нуклеотидов так же, как и пуриновый цикл, образуется из низкомолекулярных предшественников. В результате ряда последовательных реакций синтезируется уридиловая кислота (УМФ), которая, в свою очередь, является предшественником других пиримидиновых нуклеотидов - цитидиловых и тимидитиловых. Биосинтез уридиловой кислоты можно разделить на две стадии.
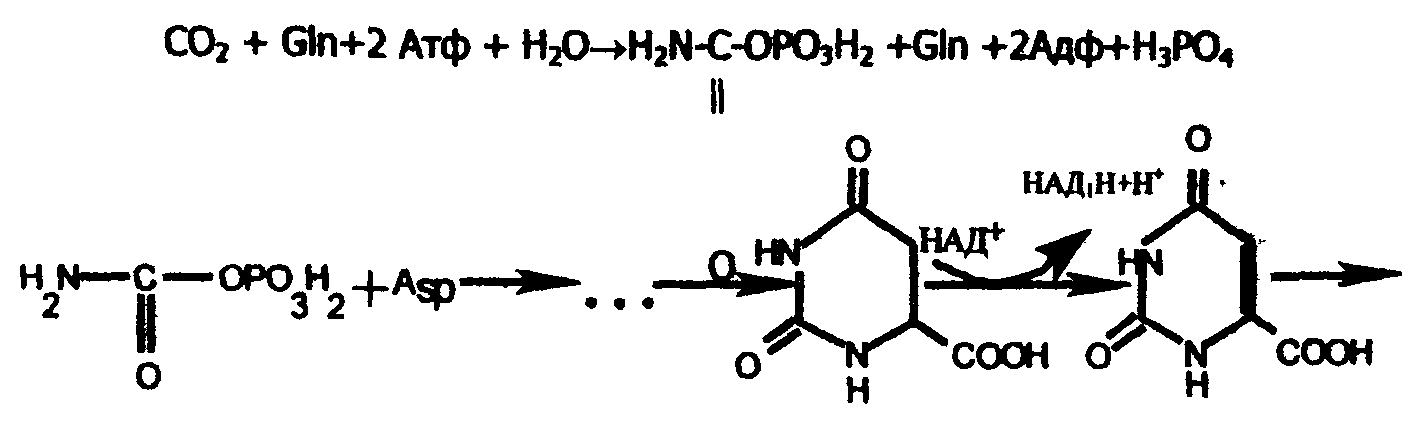
На первой стадии образуется карбомоилфосфат из глутамина и СО2 при действии цитоплазматической карбамоилфосфатсинтетазы; на второй стадии карбамоилфосфат реагирует с аспартатом и в конечном итоге образуется уридиловая кислота (УМФ):
-
карбамоилфосфат
Дигидрооротовая кислота
Оротовая кислота

Уридиновая кислота
Превращение УМФ в УДФ и УТФ осуществляется, как и в случае пуриновых нуклеотидов, путем фосфорилирования:
УМФ + АТФ ↔ УДФ + АДФ;
УДФ + АТФ ↔ УТФ + АДФ.
Предшественником цитидиловых нуклеотидов является УТФ, который превращается в ЦТФ по схеме
УТФ
+ GIn
+ АТФ ![]() ЦТФ
+ Glu
+ АДФ + Н3Р04
ЦТФ
+ Glu
+ АДФ + Н3Р04
Биосинтез дезоксирибонуклеотидов
Дезоксирибонуклеотиды - предшественники ДНК - образуются из рибонуклеотидов путем восстановления гидроксогруппы у второго углеродного атома рибозы при участии фермента - рибонуклеозидредуктазы. Субстратами фермента являются дифосфаты нуклеотидов, донором водорода служит низкомолекулярный белок тиоредоксин, содержащий две свободные SH - группы на 108 аминокислотных остатков:

Рибонуклеозиддифосфат

Дезоксирибонуклеозиддифосфат
В состав дезоксирибонуклеотидов вместо уридиловых нуклеотидов входят тимидиловые. Тимидиловая кислота (дТМФ) образуется из дезоксиуридиловой кислоты (дУМФ) путем метилирования урацила.
Непосредственные предшественники ДНК: дАТФ, дГТФ, дЦТФ, дТТФ образуются путем фосфорилирования дезоксирибонуклеозид - 5´-дифосфатов с помощью АТФ:
АТФ + дАДФ → АДФ + дАТФ;
АТФ + дГДФ → АДФ + дГТФ;
АТФ + дЦДФ →АДФ + дЦТФ;
АТФ + дТДФ → АДФ + дТТФ.
дТТФ получается также по схеме:
дУДФ → дУТФ → дУМФ → дТМФ → дТДФ → дТТФ.
Синтез дезоксирибонуклеотидов в покоящихся клетках практически не происходит и активируется на стадиях, предшествующих клеточному делению.