

Акушерство для студентов / Запоражан том 1
.pdfРозділ 4. Репродуктивна ендокринологія
винним місцем продукції IGF І в яєчниках є гранульозні клітини, хоча специфічні рецептори мають як гранульозні, так і тека клітини, що підтверджує можливий ре гуляторний механізм двокомпонентного яєчникового стероїдогенезу. Рецептори IGF ІІ знаходяться в гранульозних клітинах IGF ІІ секретується гранульозними кліти нами і посилює стероїдогенез. Хоча інсулін не виробляється в яєчниках, він може впливати на яєчниковий стероїдогенез кількома шляхами. Інсулін може взаємодія ти з рецепторами гранульозних клітин і зв’язувати рецептори IGF І у високих кон центраціях. Інсулін може регулювати рівень циркулюючого IGF І зв’язуючого про теїну (IGFBPs), який зменшений у жінок із синдромом полікістозних яєчників, інсу лінорезистентністю і гіперінсулінемією. Також ймовірно, що андрогенні прояви цих синдромів є вторинними у відповідь на дію інсуліну через IGF І рецептори в стромі яєчників.
Інгібін, активін і фолістатин
Існують докази, що інгібін, активін і фолістатин відіграють важливу роль у яєч никовому стероїдогенезі. Інгібін і активін — глікопротеїни, які складаються з двох субодиниць, з’єднаних між собою дисульфідними зв’язками. Два біоактивних інгібіни
— інгібін А та інгібін В, кожний молекулярною масою 32 000 дальтон, мають подіб ну біологічну функцію. Вони синтезуються переважно в гранульозних клітинах, яєч кових Сертолі клітинах, у жовтому тілі та плаценті. Продукція інгібіну збільшуєть ся під дією ФСГ, хоча її можуть порушувати деякі аутокринні та паракринні факто ри. Інгібін більше пригнічує виділення ФСГ, ніж ЛГ. Локальна дія інгібіну на рівні яєчників полягає у стимуляції продукції андрогенів тека клітинами і пригнічуючо му впливі на дозрівання ооцитів. Протягом менструального циклу рівень інгібіну в ранню і середню фолікулярну фазу є низьким, він підвищується через кілька днів після піка ЛГ і досягає максимуму в середині лютеїнової фази. Рівень інгібінів різко зменшується в перименопаузі та менопаузі, що може мати зв’язок з підвищенням рівня ФСГ у цьому періоді життя.
Біоактивні форми активіну мають молекулярну масу 28 000 дальтон. На рівні гіпофіза активін стимулює виділення ФСГ, на відміну від інгібіну. Паракринна/ аутокринна функція активіну включає ароматазну активність і продукцію прогесте рону, сприяння фолікулогену шляхом диференційованої дії на гранульозні клітини і попередження передчасної лютеїнізації фолікула.
Фолістатин кодується одним геном і не має структурної подібності з активіном і інгібіном. Його дія полягає у пригніченні виділення ФСГ шляхом зв’язування акти віну, і отже, зменшенні активності активіну. Більш того, фолістатин інгібує чис ленні пара та аутокринні функції активіну і прискорює темп дозрівання ооцитів шляхом впливу на гранульозні клітини.
Трансформуючий фактор росту (TGF?α ) та епідермальний фактор росту (EGF?
α ) є гомологічними β субодиниці молекул інгібіну й активіну і є потенційними ре гуляторами проліферації та диференціації гранульозних клітин.
Ітерлейкін?1 (IL 1) — поліпептидний цитокін, який секретується переважно мак рофагами. В яєчниках він утворюється тека інтерстиційними клітинами і гранульоз ними клітинами після розриву фолікула. Регуляція IL 1 на рівні яєчника первинно детермінується локальною концентрацією прогестерону. IL 1 проявляє антигона дотропну активність шляхом супресії функціональної та морфологічної лютеїнізації гранульозних клітин. Існують докази, що IL 1 може відігравати центральну роль у передовуляторній підготовці та розриві фолікула.
Простагландини. Попередником біосинтезу простагландинів (ейкозаноїдів) є ара хідонова кислота, яка вивільнюється з мембранних фосфоліпідів за допомогою фер менту ліпази. Ейкозаноїди мають широкий спектр біологічних ефектів і відіграють
129

Акушерство і гінекологія. Том 1
важливу роль у функції репродуктивної системи та фізіології яєчників. Вони конт ролюють процес початку росту фолікула шляхом сприяння посиленню кровотоку навколо фолікула і активують ФСГ рецептори гранульозних клітин преовулятор них фолікулів. Простагландини Е2 і F2α (PGЕ2 і PGF2α ) концентруються у фоліку лярній рідині преовуляторного фолікула і можуть допомагати процесу овуляції шля хом сприяння активності протеолітичних ензимів у стінці фолікула. Простагланди ни можуть допомагати регулюванню періоду життя жовтого тіла. PGЕ2 є, можливо, лютеотропним, а PGF2α чинить лютеолізуючу дію.
Простагландини також мають потенціюючий ефект на рухливість маткових труб, підсилюють стимулюючий ефект естрогенів та інгібують ефект прогестерону на кон трактильну здатність мускулатури маткових труб.
Численні простаноїди синтезуються в ендометрії: PGЕ2 і PGF2α PGІ2 і тромбок сан А2 (ТХА2). Концентрація PGЕ2 і PGF2α прогресивно зростає від проліфератив ної до секреторної фази, найвищий рівень їх спостерігається під час менструації. Ці простагландини допомагають регулювати контрактильність міометрія і регулюють процес менструації.
Яєчникові стероїди
Біохімія. Стероїди є ліпідами, що мають основну хімічну структуру — ядро, яке містить 3 шестивуглеводні кільця, приєднані до 5 вуглеводного атому (циклопен танпергідрофенантрен, або гонан) (див. рис. 4.3). Молекулярна маса гонана дорів нює 250–550 дальтон. Такі стероїди, як естрадіол і прогестерон нерозчинні у воді, але швидко розчинюються в органічних розчинниках (хлороформ, діетиловий ефір). Навпаки, стероїди, які мають приєднану сульфатну або глюкуронідну групу (кон’ю говані стероїди), такі як дегідроепіандростерон сульфат (ДГЕАС) і прегнандіол глю куронід, є водорозчинними.
Першим кроком біосинтезу яєчникових стероїдів є трансформація холестеролу до прегнонолону; з прегнонолону під дією ЛГ синтезуються за допомогою специфіч них ферментів дегідроепіандростерон, прогестерон, 17 оксипроксипрогестерон, ан дростендіон і тестостерон (див. рис. 4.4). Андростерон і тестостерон взаємодіють між собою і конвертуються в естрон і, пізніше, в естрадіол за допомогою ферменту ароматази. Цей ферментативний процес ароматизації відбувається внаслідок втрати С 19 метилової групи і утворення ароматичного кільця на С 18.
Яєчник секретує три первинних стероїди: естрадіол, прогестерон і андростенді он. Ці гормони є основними секреторними продуктами зрілого фолікула, жовтого тіла і строми яєчника відповідно (рис. 4.5, 4.6). Яєчник в деякій кількості секретує також прегненолон, 17 оксипрогестрон, тестостерон, ДГЕАС і естрон. У зв’язку з тим, що яєчники не мають ферментів 21 гідроксилази, 11 гідроксилази і 18 гідрок силази, вони не можуть синтезувати мінералокортикоїди або глюкокортикоїди. Кож ного дня яєчник секретує 0,1–0,5 мг естрадіолу; його кількість зменшується перед менструацією і зростає напередодні овуляції. Щоденна продукція прогестерону ва ріює від 0,5 мг у фолікулярну фазу до 20 мг у лютеїнову фазу. Протягом фолікуляр ної фази майже весь прогестерон синтезується наднирковими залозами і дуже мала кількість — яєчниками. Яєчник синтезує 1–2 мг андростендіону, менше ніж 1 мг ДГЕА і близько 0,1 мг тестостерону щоденно. Крім гонадного біосинтезу стероїдів, їх метаболізм здійснюється також в екстрагонадних тканинах (інтерконверсія андро стендіону і тестостерону, естрону і естрадіолу), переважно за рахунок оксидації сте роїдів у їх попередники, які мають меншу біологічну активність. Естрон також кон вертується в естрон сульфат, який має довгий період напівжиття і є найбільшим
130

Розділ 4. Репродуктивна ендокринологія
компонентом пулу циркулюючих естро генів. Хоча естрон сульфат не є біологіч но активним, фермент сульфатази в та ких тканинах, як молочна залоза і ендо метрій, може конвертувати його до ест рону, який у подальшому конвертується до естрадіолу.
Андростендіон у жировій тканині може конвертуватися в естрон (1,3 % ден ної кількості), що в більшій мірі прита манний жінкам з ожирінням (до 7 %).
Транспорт. Після виділення в кро вотік стероїди зв’язуються специфічним протеїном, — глобуліном, що зв’язує ста теві гормони (SHBG) і кортикостероїд зв’язуючим глобуліном (CBG), або з не специфічним протеїном, наприклад аль буміном. Зв’язані форми стероїдних гор монів становлять близько 95 % усього циркулюючого гормону: решту назива ють «вільним» гормоном. Наприклад, у жінок у пременопаузальному періоді близько 65 і 30 % циркулюючого тесто стерону зв’язані SHBG та альбуміном; лише 2 % є незв’язаними. Альбумін зв’я зує всі стероїди, SHBG — переважно зв’я зує дигідротестостерон, тестостерон і ес традіол (за порядком зменшення афін ності). Зв’язується з більшою афінністю до кортизолу, кортикостерону і, в меншій мірі, прогестерону. Циркулюючі рівні кожного глобуліну зростають під дією ес трогенів; рівень SHBG також збільшуєть ся при ожирінні та гіпертиреоїдизмі і зменшується під впливом андрогенів і гіпотиреоїдизму.
Метаболізм. Основними місцями ме таболізму стероїдів є печінка і, в меншій мірі, нирки. Близько 10–15 % прогестеро ну трансформується у прегнандіол — 3 глюкуронід, який є основним метаболітом прогестерону в сечі. Естрадіол і естрон конвертуються в печінці до естріолу. Ці три естрогени часто називають класични ми естрогенами, тому що вони були виді лені першими. Ці естрогени в печінці і слизовій оболонці тонкої кишки кон’югу ються у різні форми естроген сульфатів і глюкуронідів (естрон сульфат, естрадіол 17 глюкуронід, естріол 16 глюкуронід та ін.).
1
60
40
20
2
20 |
|
|
|
|
|
|
10 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
10 |
3 |
|
|
|
|
|
|
|
|
|
|
|
|
5 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
200 |
4 |
|
|
|
|
|
100 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
2 |
5 |
|
|
|
|
|
1 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
15 |
10 |
5 |
0 |
5 |
10 |
15 |
Дні від піка ЛГ
Рис. 4.5. Зміни концентрації гіпофізарних і яєчникових гормонів протягом яєчникового циклу:
1 — ЛГ, мМО/мл; 2 — ФСГ, мМО/мл; 3 — П (прогестерон), нг/мл; 4 — Е2(естрадіол), пг/мл; 5 — 17 ОГП (гідроксипрогестерон, нг/мл)
131
Акушерство і гінекологія. Том 1
1
2
3
4
5
6 |
7 |
|
|
|
13 |
|
|
14 |
8 |
9 |
8 |
10 |
|
|
|
|
|
|
11 |
|
|
12 |
|
4 |
14 |
28 |
|
Дні циклу |
|
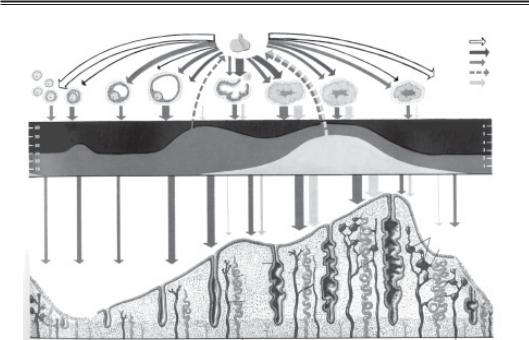
Рис. 4.6. Яєчниковий цикл:
1 — ФСГ; 2 — ЛГ; 3 — естрадіол; 4 — інгібін; 5 — прогестерон; 6 — рівень естро генів у крові; 7 — рівень прогестерону в крові; 8 — менструація; 9 — залоза; 10 — спіральна артерія; 11 — вена; 12 —базальна артерія; 13 — венозні лакуни; 14 —ана стомоз
Дія гормонів. На відміну від мембранних рецепторів протеїнових гормонів, рецеп тори стероїдних гормонів є внутрішньоклітинними. Рецептори стероїдних гормонів зв’язують специфічні класи стероїдів. Так, естрогенові рецептори зв’язують нату ральні та синтетичні естрогени, але не прогестини або андрогени. Афінність рецеп торів до стероїдів корелює з активністю останніх. Наприклад, естрогенові рецепто ри мають більшу афінність до естрадіолу, ніж до естрону та естріолу. Після зв’язу вання стероїдного гормону (С) зі своїм рецептором (Р) утворюється гормон рецеп торний комплекс (СР). Рецептори стероїдних гормонів локалізуються як у цито плазмі, так і в ядрі клітини. Вони утримуються у неактивному стані шляхом з’єднан ня з білками тіплового шоку. Зв’язуючись із гормонами, вони вивільнюють білки тіплового шоку і трансформуються так, що можуть зв’язуватися з ядерною ДНК. м РНК шляхом транскрипції утворюється з сегмента ДНК, мігрує у цитоплазму до рибосом і передає інформацію щодо синтезу нового білка. Сила сигналу для клітини залежить від концентрації гормонів, їх рецепторів, а також ступеня афінності рецеп торів до гормонів.
Естрогени стимулюють синтез як естрогенових, так і прогестеронових рецепторів у тканинах мішенях, таких як ендометрій. Прогестини пригнічують синтез як естро генових, так і прогестеронових рецепторів. Отже, вміст естрогенових та прогестеро нових рецепторів у ендометрії досягає піка в середині циклу і потім зменшується. Мітотична активність і швидкість росту ендометрія також є максимальними в сере дині циклу. Прогестини посилюють внутрішньоклітинний синтез естрадіол дегідро
132

Розділ 4. Репродуктивна ендокринологія
генази, яка конвертує більш активний естрадіол до менш активного естрону, що про гресивно зменшує естрогенну активність у клітинах мішенях.
Антиестрогени, такі як, наприклад, кломіфен або тамоксифен, зв’язують естроге нові рецептори, але ініціюють невелику транскрипцію. Отже, естрогенові рецепто ри виснажуються без нового синтезу рецепторів або дії естрогенів, що призводить до виснаження естрогенових рецепторів.
Дія гормонів на специфічні репродуктивні фактори
Яєчниковий цикл (яєчниковий гаметогенез, оогенез)
Навіть без стимуляції гонадотропінами примордіальні фолікули можуть розвива тись у первинні (преантральні) фолікули. Наявність ФСГ в овуляторному циклі призводить до зростання кількості гранульозних клітин навколо ооцита, дозрівання фолікула до антральної стадії й утворення центральної порожнини — фолікулярної печери (antrum). Без ФСГ стимуляції всі фолікули піддаються атрезії. Циклічне дозрі вання первинних фолікулів звичайно починається у періоді статевого дозрівання. Сполучна дія ФСГ і ЛГ сприяє зростанню виробки естрадіолу гранульозними кліти нами. Естрадіол стимулює ріст преантральних фолікулів, запобігає їх атрезії і поси лює дію ФСГ на гранульозні клітини. Тестостерон сприяє атрезії фолікулів і запобі гає росту преантральних фолікулів. Місцевий вміст естрогенів та андрогенів у фолі кулі впливає на шлях його розвитку: дозрівання або атрезія. Домінантний фолікул секретує велику кількість естрадіолу, що сприяє збільшенню кількості рецепторів ФСГ, зростанню кількості гранульозних клітин та їх мітотичної активності.
Підвищення рівня естрадіолу викликає ефект негативного зворотного зв’язку щодо виділення ФСГ гіпофізом і розвиток інших фолікулів припиняється (атрезія). Гранульозні клітини домінантного фолікула секретують інгібін, який також при гнічує виділення ФСГ. Ооцит оточується прозорою оболонкою (zona pellucida) та променистим вінцем (corona radiata), у фолікулі накопичується рідина, яка містить естрогени, андрогени та протеїни. Після досягнення критичної маси гранульозних клітин у фолікулі виникає печера (antrum) і ооцит переміщується в бік фолікула (рис.4.7). За даними ультразвукового дослідження, домінантний фолікул досягає діа метра в середньому 19 мм (18–25 мм), а його об’єм зростає до 3,8 мл (3,1–8,2 мл). Близько ооцита гранульозні клітини нагромаджуються і утворюють яйценосний гор бок (cumulus oophorus). Під час проліферації гранульозних клітин на їх поверхні ут ворюються рецептори до ЛГ. При зв’язуванні молекул ЛГ зі специфічними рецепто рами проліферація гранульозних клітин припиняється і вони починають секретува ти прогестерон.
Близько 80 % усієї кількості естрадіолу (500 мкг) продукується домінантним фо лікулом напередодні овуляції ( за 12–24 год). Зростання рівня естрадіолу і, в меншій мірі, прогестерону стимулює секрецію ЛГ гіпофізом (позитивний зворотний зв’я зок). Пік ЛГ у середині циклу ініціює процес овуляції — розриву стінки зрілого фолікула і виходу ооцита в черевну порожнину і в подальшому в просвіт маткової труби. Овуляція виникає приблизно через 24 год після досягнення піка секреції
133
Акушерство і гінекологія. Том 1
1
2
3
4
5
6
7
8
9 10
11
13 12
14
15
А
20
19
18
17
25
28
29 2726
30
31
32
33
34
35
36
16
Б
21
22
23
24
Г |
В |
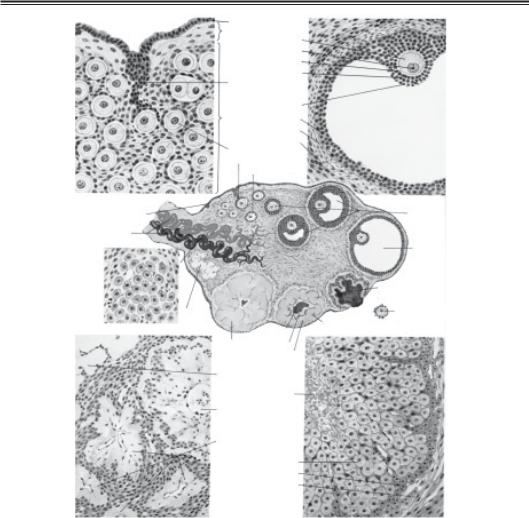
Рис. 4.7. Анатомія і фізіологія яєчника (А — яєчник новонародженої дівчин ки; Б — зрілий фолікул; В — жовте тіло: Г — яєчник жінки в постменопаузі):
1 — поверхневий епітелій яєчника; 2 — білкова оболонка (tunica albuginea); 3 — яйцеклітина; 4 — ядро; 5 — блискуча зона (zona pellucida); 6 — променистий вінець (corona radiata); 7 — тяжі епітеліальних клітин, що мігрують донизу і оточують яй цеклітину; 8 — яйценосний горбок (cumulus oophorus); 9 — кора яєчника (cortex); 10 — гранульозна оболонка фолікула; 11 — внутрішня тека оболонка фолікула (thеса interna); 12 — зовнішня тека оболонка фолікула (theca externa); 13 — при мордіальний фолікул; 14 — епітеліальний тяж; 15 — первинний фолікул; 16 — по рожнина зрілого фолікула, наповнена фолікулярною рідиною (antrum); 17 — біле тіло (corpus albicans); 18 — гілюсні клітини; 19 — кровоносні судини, що входять у ворота яєчника; 20 — поверхневий епітелій; 21 — фолікул, що розвивається; 22 — зрілий фолікул; 23 — геморагічне жовте тіло (розірваний фолікул); 24 — звільнена яйцеклітина; 25 — раннє жовте тіло; 26 — лютеїнові клітини; 27 — фібрин; 28 — кров’яний згусток; 29 — зріле жовте тіло; 30 — біле тіло; 31 — фолікулярна порож нина (зі згустком і фібрином); 32 — фолікул у стадії ранньої атрезії; 33 — пізні ат ретичні фолікули; 34 — гранульозні лютеїнові клітини; 35 — тека лютеїнові кліти ни; 36 —зовнішня тека оболонка
134

Розділ 4. Репродуктивна ендокринологія
естрадіолу, через 32 год після початку зростання рівня ЛГ і між 12 і 16 год після досягнення піка ЛГ у сироватці крові.
Хвиля ЛГ у середині циклу ініціює завершення метафази першого ділення. Ооцит вступає у метафазу ІІ ділення і утворюється перше полярне тільце. Завершення мейозу і утворення другого полярного тільця відбуваються тоді, коли яйцеклітина пенетрується сперматозоїдом. ЛГ стимулює синтез простагландинів і протеолітич них ензимів. Плазмін під дією ФСГ сприяє процесу виштовхування ооцита і куму люса під час розриву фолікула. Звільнений ооцит потрапляє в порожнину маткової труби за допомогою рухів її торочок і далі в матку.
Після овуляції кількість фолікулярної рідини, діаметр і об’єм фолікула зменшу ються. Фолікулярна стінка скручується, місце розриву фолікула — стигма овуляції
— закривається фібрином. Якщо запліднення не відбувається, утворюється жовте тіло менструації (corpus luteum). Під дією ЛГ жовте тіло вступає у стадію розквіту і секретує прогестерон (20 мкг/добу), а також естрадіол. Прогестерон забезпечує фазу секреції в менструальному циклі, сприяє підготовчим змінам у слизовій оболонці матки, спрямованим на здійснення імплантації, забезпечує фізіологічний перебіг вагіт ності. Високий рівень ЛГ, а також ХГЛ необхідні для підтримки секреторної функції жовтого тіла. Стадія розквіту жовтого тіла триває 12–14 днів; діаметр жовтого тіла менструації досягає 1,5–2 см. Якщо імплантації не відбувається, жовте тіло підлягає зворотному розвитку. Його клітини підлягають жировій дегенерації, і утворюється біле тіло (сorpus albicans), яке з часом абсорбується стромою яєчника.
Рівень прогестерону рівномірно зростає у сироватці крові після овуляції і дося гає плато через 1 тиж і потім зменшується, якщо не виникає імплантації. Зростаючі рівні естрогенів та прогестерону за механізмом негативного зворотного зв’язку при гнічують секрецію ФСГ і ЛГ. Естрадіол інгібує перважно ФСГ (негативний зворот ний зв’язок), прогестерон — ЛГ. Продукція прогестерону жовтим тілом спричинює місцевий лютеолітичний ефект. Вважають, що зростання рівня прогестерону в яєч нику запобігає дозріванню фолікула в цьому яєчнику в наступному циклі. Після закінчення лютеолізу і зменшення рівнів естрадіолу і прогестерону виникає слаб ший зворотний зв’язок. Перед початком менструації починають зростати рівні ФСГ і ЛГ для подальшої стимуляції росту фолікулів у наступному яєчниковому циклі.
Естрадіол і прогестерон чинять пряму пригнічуючу дію на синтез і секрецію гіпофізарних гонадотропінів, а також на вивільнення ГнРГ, змінюючи частоту та амплітуду його імпульсів. Цей стероїдний вплив здійснюється за допомогою нейро трансмітерів (допаміну та норепінефрину) і нейромодуляторів (β ендорфінів) в ар куатних ядрах гіпоталамуса. В ранню фолікулярну фазу пульсація ЛГ відбувається кожні 90 хв, припиняється під час сну і зростає в середній та пізній фолікулярній фазі (1 імпульс за 60 хв вдень і вночі) і уповільнюється до 1 імпульсу за 90 хв у ранню лютеїнову фазу і 1 імпульсу за 3 год у пізню лютеїнову фазу. Подібні зміни пульсації ФСГ у периферичній крові не відбуваються, можливо, у зв’язку з його довшим періодом напівжиття.
Зростання частоти імпульсів ЛГ у пізню фолікулярну фазу є важливим для сти муляції секреції естрадіолу. Близько 80 % пульсуючих викидів ЛГ відбуваються після підйому рівня естрогенів у периферичній крові (позитивний зворотний зв’язок). Пульсація ЛГ в середню і пізню лютеїнову фази стимулює пульсуючу продукцію прогестерону. Зменшення частоти імпульсів ЛГ протягом середньої лютеїнової фази може відбуватися внаслідок негативної зворотної дії прогестерону і естрадіолу. Змен шення частоти секреції ЛГ пов’язують із впливом прогестерону на вивільнення ГнРГ гіпоталамусом. Медіаторами цього процесу є, можливо, β ендорфіни. В свою чергу зменшення амплітуди імпульсів ЛГ пов’язують із негативною зворотною дією про гестерону на гіпофіз.
135

Акушерство і гінекологія. Том 1
Екскреція ФСГ і ЛГ із сечею коливається від 1 до 10 МО/добу, а їх рівень у сироватці крові — від 1 до 100 мМО/мл. Екскреція естрогенів (естрону, естрадіолу і
естріолу) є найнижчою протягом ранньої фолікулярної фази, досягає максимуму без посередньо перед піком ЛГ, після чого швидко знижується і знову підвищується в лютеїнову фазу, наприкінці якої також знижується. Зростання рівня цих естрогенів у лютеїнову фазу має меншу амплітуду, але більшу тривалість, ніж протягом передову ляторного піка. Пік екскреції естрогенів із сечею становить 50–75 мкг/добу. Сироват ковий рівень естрадіолу зростає з менш ніж 50 пг/мл у ранню фолікулярну фазу до 200–500 пг/мл у середині циклу з підвищенням у лютеїнову фазу до 100–150 пг/мл.
Основним метаболітом прогестерону, який екскретується з сечею, є прегнандіол. Рівень прегнандіолу не перевищує 0,9 мкг/добу перед овуляцією і вищий 1 мкг/добу (3–4 мкг/добу) після овуляції. Рівень прогестерону у сироватці крові менший 1 нг/мл перед овуляцією і в середині лютеїнової фази досягає 10–20 нг/мл. За 4– 6 днів до початку менструації рівні естрадіолу, прогестерону і 17 гідроксипрогестеро ну починають знижуватись. У середині циклу спочатку підвищується рівень естрадіо лу. Після досягнення піка естрадіолу виникає раптовий підйом хвилі ЛГ і ФСГ. Сек реція ЛГ досягає піка через 18 год і утримується (плато) протягом 14 год, а потім знижується. Середня тривалість хвилі ЛГ дорівнює 24 год. За 12 год до початку хвилі ЛГ зростає рівень прогестерону і 17 гідроксипрогестерону. Разом з піком ЛГ спостері гається зниження концентрації естрадіолу і подальше зростання рівня прогестерону. Таке зрушення стероїдогенезу на користь прогестерону замість продукції естрадіолу відбувається внаслідок лютеїнізації гранульозних клітин фолікула під дією ЛГ.
Рівні інших гормонів протягом яєчникового циклу також підлягають деяким змінам. Сироваткові рівні андростендіону і тестостерону незначно коливаються протягом цик лу, хоча їх середні рівні є вищими протягом фолікулярної фази порівняно з лютеїно вою. Рівень ТТГ є відносно стабільним, тимчасом як вміст адренокортикотропного гормону (адренокортикотропіну, АКТГ) і гормону росту мають передовуляторний пік, а рівень пролактину дещо підвищується у лютеїнову фазу циклу.
Якщо відбувається запліднення, продукція прогестерону жовтим тілом триває під стимулювальним впливом ХГЛ, який секретується синцитіотрофобластом і ут ворюється жовте тіло вагітності. Жовте тіло досягає в діаметрі 1,5–2 см і зазнає зворотного розвитку через 11–12 тиж.
Тканини мішені. Менструальний цикл
Довжина (тривалість) менструального циклу. Середній вік менархе становить 12– 13 років, а середній вік менопаузи — близько 51 року. Отже, менструація у жінки відбувається понад 38 років. Довжина менструального циклу коливається у різних жінок і у однієї і тієї самої жінки протягом різних періодів її життя. Нерегулярний менструальний цикл частіше спостерігається протягом 2 років після менархе і 3 років перед менопаузою, коли найбільш частими є ановуляторні цикли. Протягом цих пе ріодів життя можуть спостерігатись як скорочення, так і продовження (частіше) дов жини менструального циклу.
Довжина менструального циклу є найменш варіабельною у віці між 20 та 40 ро ками. Протягом цього періоду відбувається поступове зменшення довжини циклу. Фолікулярна фаза визначається як інтервал між початком (1 м днем) менструації та днем, коли починається зменшення пікового рівня ЛГ. Прийнято вважати, що серед ня тривалість менструального циклу дорівнює 28±7 днів. Короткі менструальні цик ли (<21 дня) називаються поліменореєю, довгі (>35 днів) — олігоменореєю. Серед ня тривалість менструальної кровотечі дорівнює 4±2 дні.
136

Розділ 4. Репродуктивна ендокринологія
Зміни ендометрія в менструальному (ендометріальному) циклі
Ендометрій має два основних шари: базальний і функціональний. Зміни ендомет рія в менструальному циклі відбуваються у його функціональному шарі, який част ково десквамується під час менструації (див. рис. 4.6).
Після менструації товщина ендометрія дорівнює 1–2 мм і він складається пере важно з базального шару і частини функціонального шару. Під дією естрогенів у фо лікулярну фазу яєчникового циклу в ендометрії починається фаза проліферації. Про ліферація відбувається у функціональному шарі ендометрія за рахунок збільшення кількості залозистих і стромальних клітин. У клітинах спостерігаються мітози. В пізню фолікулярну (проліферативну) фазу залози стають більш звивистими; під дією естрогенів у передовуляційний період їх клітини підлягають псевдостратифікації. Відразу після овуляції в клітинах залоз ендометрія з’являються під’ядерні вакуолі, які містять глікоген. Під’ядерна вакуолізація клітин є свідченням дії прогестерону, а не овуляції. Під дією прогестерону в ранню лютеїнову фазу в ендометрії починаєть ся фаза секреції і глікогенвміщуючі вакуолі виштовхуються у просвіт залоз. Зреш тою вміст залоз потрапляє у порожнину ендометрія, де глікоген зберігається для підтримання життєдіяльності бластоцисти, яка з’являється у порожнині матки через 3,5 дні після фертилізації.
У середині лютеїнової (секреторної) фази залози ще більше скручуються, відбу вається набряк і васкуляризація строми. Протягом секреторної фази в ендометрії ви робляється кілька специфічних протеїнів: плацентарні протеїни 14 і 12 (РР14 і РР12), ендометріальний протеїн, асоційований з вагітністю (РЕР) і α 2 асоційований з вагіт ністю ендометріальний протеїн (глікоделін), а також пептидні гормони, фактори ро сту і простагландини. РР12 називають ще α матковим протеїном і хоріонічним α 2 глобуліном. Циркулюючий рівень РР12 корелює з рівнем прогестерону. Всі ці суб станції відіграють певну роль у процесі децидуалізації ендометрія. Якщо в пізню лю теїнову (секреторну) фазу імплантації не відбувається, ХГЛ не продукується, жовте тіло підлягає зворотному розвитку, залози ендометрія спадаються і фрагментуються. Відбувається інфільтрація залоз і строми лейкоцитами і макрофагами, аутоліз і дес квамація функціонального шару ендометрія (фаза менструації).
До інших органів та клітин?мішеней, які мають рецептори до яєчникових стерої дних гормонів і можуть відповідати на їх дію, належать молочні залози, жирова, м’я зова, кісткова тканина, шкіра та її придатки (потові та сальні залози, волосяні фолі кули), ендотелій судин, кардіоміоцити, мозок та ін.
Контрольні питання
1.Перерахуйте прямі та зворотні зв’язки різних рівнів регуляції репродуктивної системи.
2.Охарактеризуйте ендокринні основи яєчникового циклу.
3.Які зміни органів мішеней відбуваються в яєчниковому циклі?
4.Що таке примордіальний, преантральний, антральний фолікул?
5.Які гормони є необхідними для накопичення гранульозних клітин
іпрогресивного росту фолікула?
137

Акушерство і гінекологія. Том 1
6.Який гормон дає сигнал для розвитку нового фолікула?
7.Яка функція лютеїнізуючого гормону у фолікулярній фазі циклу?
8.Як відбувається стероїдогенез у яєчниках?
9.Які гормони гальмують дозрівання фолікула?
10.Завдяки яким механізмам відбувається атрезія недомінантних фолікулів?
11.Назвіть предиктори овуляції.
12.Що необхідне для повноцінного розвитку жовтого тіла?
13.Коли відбувається імплантація?
14.Що таке «недостатність лютеїнової фази менструального циклу»?
138