

Акушерство для студентов / Запоражан том 1
.pdfРозділ 5. Репродуктивна генетика...
Підтримуючі клітини Сертолі походять з клітин поверхневого епітелію яєчка, подібно до фолікулярних клітин яєчника.
Невдовзі перед статевим дозріванням у статевих шнурах виникає просвіт, і вони перетворюються у сім’яні канальці. В цей час первинні статеві клітини дають поча ток сперматогоніям, серед яких є 2 типи: сперматогонії типу А і сперматогонії типу Б. Сперматогонії типу А поділяються мітозом і забезпечують резерв стовбурових клітин, сперматогонії типу Б дають початок первинним сперматоцитам. За нормаль них умов деякі клітини типу А виходять з популяції стовбурових клітин, щоб запо чаткувати подальші генерації сперматогоній, кожна з яких є більш диференційова ною порівняно з попередньою генерацією. Після завершення останнього поділу клітин типу А формуються сперматогонії типу Б, які після серії мітозів перетворю ються у первинні сперматоцити. Первинні сперматоцити вступають у продовжену профазу (22 дні), після якої настає швидке завершення мейозу І з утворенням вто? ринних сперматоцитів. Вторинні сперматоцити відразу починають другий мейотич ний поділ, внаслідок чого утворюються гаплоїдні сперматиди. Протягом цих про цесів, починаючи від моменту, коли клітини типу А залишають популяцію стовбу рових клітин, і до утворення сперматид, цитокінез не завершується і клітини по слідовних генерацій залишаються зв’язаними цитоплазматичними містками. Отже, нащадки однієї сперматогонії типу А формують кластер статевих клітин, що зберіга ють контакт між собою впродовж усієї диференціації. Крім того, сперматогонії та сперматиди протягом свого розвитку занурені у глибокі кишені з клітин Сертолі. Клітини Сертолі забезпечують підтримку для дозріваючих статевих клітин, їх за хист і живлення, створюють умови для вивільнення зрілих сперматозоїдів.
Сперміогенез — низка змін, внаслідок яких сперматиди перетворюються на спер матозоїди. У людини процес перетворення сперматогонії на зрілий сперматозоїд три ває 64 доби. Сперміогенез включає такі фази:
1)формування акросоми, яка вкриває половину поверхні ядра і містить ензими, що допомагають пенетрації в яйцеклітину через її мікрооточення під час запліднення;
2)конденсацію ядра;
3)формування шийки, середньої частини і хвоста;
4)відторгнення більшої частини цитоплазми.
Сформовані сперматозоїди направляються у просвіт сім’яних канальців. Із сім’я них канальців шляхом скорочення елементів їх стінки сперматозоїди просуваються у канальці придатка яєчка. Спочатку сперматозоїди є малорухливими і набувають активної рухливості у придатку яєчка.
Клінічні кореляції. Аномальні гамети. У людини, як і в більшості ссавців, один фо лікул яєчника інколи може містити два або три первинні ооцити. Такі ооцити мо жуть дати початок двійні або трійні, але зазвичай вони дегенерують, не досягнувши стадії зрілості. У рідкісних випадках один ооцит може містити два або три ядра. Такі багатоядерні ооцити також гинуть, не досягнувши зрілості.
На відміну від ооцитів, аномалії сперматозоїдів зустрічаються часто (близько 10 % сперматозоїдів мають дефекти). Голівка, хвіст, сперматозоїди можуть бути ма ленькими, гігантськими або з’єднаними. Сперматозоїди з відхиленнями у морфо логії не мають нормальної рухливості і зазвичай не здатні до запліднення ооцитів.
Запліднення
Запліднення — це процес злиття чоловічої та жіночої гамет. Запліднення відбу вається в ампулярній частині маткової труби.
Ооцит, який виштовхується з фолікула разом з оточуючими його клітинами гра нульози ділянки яйценосного горбка, потрапляє в ампулу маткової труби. Деякі
149

Акушерство і гінекологія. Том 1
клітини яйценосного горбка після цього переорієнтовуються навколо прозорої зони й утворюють променистий вінець, або корону (corona radiata). У момент викиду ооци та, оточеного клітинами яйценосного горбка, з яєчника (овуляція) завершується пер ший мейотичний поділ і вторинний ооцит починає другий мейотичний поділ.
Сперматозоїди зберігають здатність до запліднення в жіночих статевих шляхах протягом кількох діб. Вони швидко проникають із піхви в шийку матки, матку і маткові труби, що зумовлено руховою активністю хвостів сперматозоїдів у шийко вому слизу та скороченнями мускулатури матки і маткової труби. Для спермато зоїдів, які опиняються у матковій трубі, перешийок служить резервуаром, а рух із цієї ділянки до ампули є синхронізованим процесом.
Сперматозоїди, які щойно потрапили у жіночі статеві шляхи, не здатні до заплід нення ооцита. Для набуття здатності до запліднення їм треба здійснити капацита?
цію і акросомну реакцію.
Капацитація — це хімічна модифікація (кондиціонування) поверхні спермато зоїдів і сім’яної рідини у жіночих статевих шляхах. У людини цей процес триває близько 7 год. Суть капацитації, яка відбувається в матковій трубі, полягає у взає модії між сперматозоїдом і поверхнею слизової оболонки маткової труби. При цьо му глікопротеїнове покриття і білки сім’яної рідини видаляються з плазматичної мембрани акросомної ділянки сперматозоїда. Тільки капацитовані сперматозоїди можуть пройти крізь променистий вінець ооцита і здійснити акросомну реакцію.
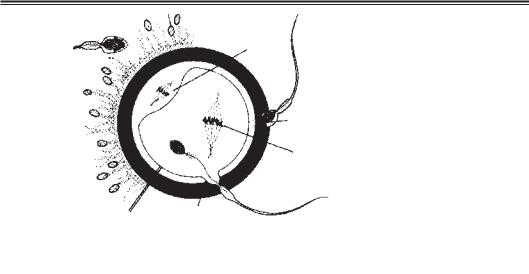
Акросомна реакція відбувається після зв’язування сперматозоїда з прозорою зо ною і зумовлюється білками останньої. Суть акросомної реакції полягає у вивіль ненні ензимів, необхідних для пенетрації прозорої зони — акрозину і трипсиноподі бних речовин (рис. 5.6).
Запліднення складається з трьох фаз, або стадій:
1)пенетрація променистого вінця;
2)пенетрація прозорої зони;
3)злиття мембран ооцита і сперматозоїда.
Устадії 1 із 200–300 млн сперматозоїдів, які потрапляють у жіночі статеві шляхи при сім’явипорскуванні, лише 300–500 досягають місця запліднення і тільки один з них запліднює яйцеклітину; інші сперматозоїди, можливо, допомагають йому про никнути крізь бар’єри, що оточують ооцит. Капацитовані сперматозоїди легко про никають через клітини променистого вінця.
Устадії 2 прозора зона — глікопротеїнова оболонка, що оточує ооцит, забезпечує зв’язування сперматозоїдів та індукує акросомну реакцію. Зв’язування спермато зоїдів з прозорою зоною індукується лігандом ZP3, білком прозорої зони і рецепто рами сперматозоїда. Вивільнення акросомних ензимів (акрозину) дозволяє сперма тозоїду проникнути крізь прозору зону і вступити в контакт з плазматичною мемб раною ооцита. Після взаємодії голівки сперматозоїда з поверхнею ооцита проникність прозорої зони змінюється. Результатом цієї взаємодії є секреція лізосомних фер ментів кортикальних гранул, що містяться під плазмолемою ооцита. Ці ферменти викликають зміну властивостей прозорої зони (зона — реакція), що запобігає пенет рації інших сперматозоїдів та інактивує видоспецифічні сайти рецепторів спермато зоїдів на поверхні прозорої зони. У прозору зону можуть заглиблюватися багато сперматозоїдів, але лише один із них проникає в ооцит.
Устадії 3 первинна адгезія сперматозоїда з ооцитом частково опосередковується взаємодією інтегринів поверхні ооцита та їхніх лігандів на сперматозоїді. Відразу після адгезії відбувається злиття цитоплазматичних мембран сперматозоїда та яйце клітини. Оскільки плазматична мембрана акросомної ділянки сперматозоїда зникає
врезультаті акросомної реакції, зливаються мембрана ооцита і мембрана задньої частини голівки сперматозоїда. У людини в цитоплазму ооцита входять голівка і
150
Розділ 5. Репродуктивна генетика...
|
Фаза 1 |
|
Полярне тільце |
|
|
Фаза 2 |
|
|
|
|
|
у стадії мейозуІІ |
|
|
|
|
|
|
|
|
Розчинення внутріш |
|||||
|
|
|
|
|
||||
|
Акросома |
|
|
ньої мембрани акросоми |
||||
|
|
|
|
|
|
|
||
|
Ядро |
|
|
Вторинний ооцит |
||||
сперматозоїда |
|
|
у стадії мейозуІІ |
|||||
|
|
Плазматична |
|
|
|
|
|
|
|
|
мембрана |
Злиття клітинних |
|
|
|
|
|
|
|
|
мембран ооцита |
|
|
|
|
|
|
|
|
і сперматозоїда |
|
|
|
|
|
|
|
|
Фаза 3 |
|
|
|
||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
Рис. 5.6. Три фази пенетрації яйцеклітини сперматозоїдом
хвіст сперматозоїда, тимчасом як плазмолема сперматозоїда залишається фіксова ною до поверхні ооцита. На проникнення сперматозоїда ооцит реагує трьома проце сами:
1.Кортикальна реакція та зона?реакція. Внаслідок викидання з ооцита корти кальних гранул, у яких містяться лізосомні ферменти, мембрана ооцита стає непро никною для інших сперматозоїдів і прозора зона змінює свою структуру та склад, запобігаючи зв’язуванню і проникненню інших сперматозоїдів. Цей механізм запобі гає поліспермії (проникненню в ооцит більш ніж одного сперматозоїда).
2.Поновлення другого мейотичного поділу. Одразу після входження сперматозоїда ооцит завершує свій другий мейотичний поділ. Одна з дочірніх клітин майже не отримує цитоплазми — друге полярне тільце, інша — є дефінітивним ооцитом. Хро мосоми дефінітивного ооцита (22+Х) утворюють пухирчасте ядро — жіночий про? нуклеус (рис. 5.7).
3.Метаболічна активація яйцеклітини. Фактор активації, можливо, вноситься сперматозоїдом. Активація після злиття ооцита зі сперматозоїдом включає в себе первинні клітинні і молекулярні процеси, пов’язані з раннім ембріогенезом.
Після проникнення через оболонки ооцита сперматозоїд рухається вперед, до досягнення тісної близькості з жіночим пронуклеусом. Ядро сперматозоїда набухає
іутворює чоловічий пронуклеус, тимчасом як хвіст відокремлюється і дегенерує. Морфологічно чоловічий і жіночий пронуклеуси розрізнити неможливо. Вони всту пають у тісний контакт, втрачають ядерні оболонки і реплікують свої ДНК. Одразу після синтезу ДНК хромосоми утворюють веретено поділу і готуються до мітозу. Материнські (23) і батьківські (23) двохроматидні хромосоми розщеплюються по здовжньо в ділянці центромери. Сестринські хроматиди рухаються до протилежних полюсів, забезпечуючи кожну клітину двоклітинного зародка нормальною диплоїд ною кількістю хромосом і ДНК. Коли сестринські хроматиди рухаються до проти лежних полюсів, на поверхні клітини утворюється глибока борозна, яка з часом розділяє цитоплазму на 2 частини.
151

Акушерство і гінекологія. Том 1
Променистий |
Прозора зона |
Перивітелиновий |
Чоловічий |
вінець |
|
простір |
пронуклеус |
Веретено другого |
Жіночий |
Полярні тільця |
Центросома |
поділу дозрівання |
пронуклеус |
|
|
Двоклітинна
стадія
Рис. 5.7. Ооцит безпосередньо після овуляції
Отже, основні результати запліднення включають такі процеси:
1)відновлення диплоїдного набору хромосом, половина з яких походить від батька, половина — від матері. Утворена зигота містить нову комбінацію хромосом, що відрізняється від генотипу батьків;
2)визначення статі нового індивіда. Сперматозоїд, що несе Х хромосому, обу мовлює розвиток ембріона жіночої статі (ХХ), а сперматозоїд, що несе Y хромосо му, ембріона чоловічої статі (XY);
3)початок дроблення. Якщо запліднення не відбувається, ооцит зазвичай дегене рує через 24 год після овуляції.
Клінічні кореляції. Проблема неплідності торкається 15–30 % подружніх пар. Чо ловіча неплідність зумовлена аномаліями кількості і якості сперматозоїдів. У нормі еякулят має об’єм 3–4 мл і містить 100 млн сперматозоїдів. Якщо кількість сперма тозоїдів < 50 млн в еякуляті (або < 20 млн в 1 мл сперми), має місце олігозооспермія
ічоловіча неплідність. Жіноча неплідність може бути зумовлена анатомічними чин никами (непрохідність маткових труб та ін.), відсутністю овуляції (ановуляцією), несприятливим впливом цервікального слизу на сперматозоїди; імунітетом до спер матозоїдів та іншими причинами.
При екстракорпоральному заплідненні, або фертилізації in vitro (IVF), ріст фолі кулів у яєчниках стимулюють введенням гонадотропінів; забирають ооцити перед овуляцією, вміщують їх у культуральне середовище і додають сперматозоїди. Заплід нені яйцеклітини культивують до 8 клітинної стадії, а потім імплантують у матку. Ембріони доімплантаційних стадій є нечутливими до дії тератогенних чинників, тому ризик вад розвитку при здійсненні маніпуляцій при екстракорпоральному за плідненні не збільшується. До інших методів екстракорпорального запліднення на лежать пересадка гамет у маткову трубу (GIFT) або пересадка зиготи у маткову трубу (ZIFT). При олігозооспермії або азооспермії виконують ін’єкцію єдиного спер
152

Розділ 5. Репродуктивна генетика...
матозоїда в цитоплазму яйцеклітини — інтрацитоплазматичну ін’єкцію сперматозої да (ICSI).
Дроблення
Після досягнення зародком двоклітинної стадії він здійснює серію мітотичних поділів (процес дроблення), що приводить до збільшення кількості клітин. Ці кліти ни — бластомери — зменшуються з кожним поділом дроблення і до восьмиклітинної стадії формують нещільний конгломерат. Після третього поділу дроблення бласто мери максимально збільшують площу міжклітинних контактів, формуючи компакт ну клітинну масу, яка утримується разом завдяки щільним контактам. Процес ком? пактизації призводить до відокремлення зовнішніх клітин від внутрішніх, які спо лучаються між собою щілинними контактами (нексусами). Приблизно через 3 год після запліднення клітини компактизованого ембріона знову діляться, утворю ючи 16 клітинну морулу (ягода шовковиці). Внутрішні клітини морули утворюють
внутрішню клітинну масу, а зовнішні клітини — зовнішню клітинну масу. Внутріш ня клітинна маса дає початок тканинам ембріона, а зовнішня клітинна маса утворює
трофобласт, з якого формується плацента.
Бластоциста
Коли морула потрапляє в порожнину матки, через прозору зону з мікрооточення ембріона починає просочуватися рідина, яка накопичується у міжклітинних просто рах внутрішньої клітинної маси. Поступово міжклітинні простори зливаються і фор мують єдину порожнину — бластоцель. Тимчасом ембріон знаходиться у стадії бла? стоцисти (рис. 5.8). Клітини внутрішньої клітинної маси утворюють ембріобласт і локалізовані на одному полюсі, тимчасом як клітини зовнішньої клітинної маси, або трофобласт, стають плоскими і утворюють стінку бластоцисти. Прозора зона зни кає, що дає можливість бластоцисті здійснити імплантацію.
Клітини трофобласта, які локалізуються над ембріобластним полюсом бластоци сти, близько шостого дня починають вростати в епітелій ендометрія — слизової обо лонки матки. Прикріплення та інвазія трофобласта забезпечуються дією інтегринів, що виділяються клітинами трофобласта, а також ламініном і фібронектином міжклітинного матриксу ендометрія. Інтегринові рецептори для ламініну сприяють адгезії, тимчасом як рецептори для фібронектину стимулюють міграцію. Взаємодія цих молекул відбувається і при передачі інформації, що регулює диференціацію тро фобласта. Таким чином, імплантація є результатом комплексних взаємодій трофо бласта й ендометрія. До кінця першого тижня розвитку зародок людини проходить стадію морули, бластоцисти і починає імплантацію у слизову оболонку матки.
Отже, імплантація відбувається наприкінці першого тижня розвитку.
Клінічні кореляції. Аномальні зародки зазвичай гинуть через 2–3 тиж після заплід нення, тому їх частоту визначити важко. Вважають, що близько 50 % вагітностей закінчуються мимовільними викиднями і половина з них зумовлені хромосомними аномаліями. Ці викидні є наслідком так званих «пренатальних фільтрів», що відбра ковують аномальні ембріони і тим самим зменшують кількість індивідів із вродже ними вадами до 2–3 % замість 12 %. При комбінації методів екстракорпорального запліднення і полімеразної ланцюгової реакції (ПЛР) проводиться молекулярний скринінг ембріонів з генетичними аномаліями (преімплантаційна діагностика). Для цього достатньо виділити один бластомер з ембріона ранньої стадії розвитку і реплі фікувати його ДНК для подальшого дослідження. Програма «Геном людини» зай мається вивченням зв’язку специфічних генів із різними клінічними синдромами.
153

Акушерство і гінекологія. Том 1
|
|
Лакуни трофобласта |
Розширені кровоносні судини |
|
||||||
А |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Синцитіотрофобласт |
|||
|
|
|
|
|
|
|
Цитотрофобласт |
|||
|
|
|
|
|
|
|
Амніотична порожнина |
|||
|
|
|
|
|
|
|
Епібласт |
|
||
|
|
|
|
|
|
|
Гіпобласт |
|
|
|
Екзоцеломічна порожнина |
Згусток |
Екзоцеломічна мембрана (Гейзера) |
||||||||
(первинний жовтковий мішок) |
фібрину |
|
|
|
|
|||||
Б |
|
|
|
|
Трофобластні лакуни |
Синцитіотрофобласт |
||||
|
|
|
|
|
|
|||||
|
|
Синцитіо |
Амніотична |
|
Війки |
|
Амніон |
|||
Епітелій |
порожнина |
|
|
Біламінарний |
||||||
матки |
трофобласт |
|
|
|
|
|
||||
|
|
|
|
|
ембріональ |
|||||
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
ний диск |
|
Внутрішня |
|
|
|
|
Епібласт |
|
|
Первинний |
||
|
|
|
|
Цитотро |
жовтковий |
|||||
клітинна |
|
|
|
|
||||||
|
|
|
|
|
|
мішок |
||||
маса |
|
|
|
|
Гіпобласт |
трофобласт |
||||
Цитотро |
Зародкова |
Екзоцеломна |
||||||||
Гіпобласт |
|
Зародкова |
|
|||||||
фобласт |
порожнина |
|
|
мембрана |
||||||
|
порожнина |
|
||||||||
|
|
|
|
|
|
|
|
|||
Рис. 5.8. Бластоциста: А — бластоциста людини 9 добового розвитку; Б — нідація через 7–10 днів після запліднення
Двошаровий зародковий диск
На другому тижні розвитку (8 ма доба) бластоциста частково занурена у строму ендометрія. В ділянці над ембріобластом трофобласт диференціюється на два шари: внутрішній шар одноядерних клітин — цитотрофобласт, зовнішню багатоядерну масу без чітких клітинних меж — синцитіотрофобласт. Фігури мітозу спростеріга ються лише у цитотрофобласті; у синцитіотрофобласті вони відсутні. Клітини ци тотрофобласта поділяються і мігрують у синцитіотрофобласт, там зливаються і втра чають свої індивідуальні клітинні оболонки. Клітини ембріобласта також диферен ціюються на два шари: шар маленьких кубоїдальних клітин, які прилягають до по рожнини бластоцисти, — гіпобласт і шар високих циліндричних клітин, які приля гають до амніотичної порожнини, — епібласт (див. рис. 5.8). Разом ці два шари ембріобласта утворюють плоский двошаровий зародковий диск. Водночас усередині епібласта утворюється маленька порожнина, яка з часом розширюється і утворює амніотичну порожнину. Клітини епібласта, прилеглі до цитотрофобласта (амніобла сти), разом з іншими клітинами епібласта вистеляють амніотичну порожнину. Стро ма ендометрія навколо місця імплантації характеризується набряком і вираженою
154

Розділ 5. Репродуктивна генетика...
васкуляризацією; великі розгалужені залози продукують значну кількість глікогену і слизу.
На 9–10 ту добу бластоциста глибше занурюється в ендометрій. Трофобласт на ембріональному полюсі розвивається найбільш інтенсивно, в його синцитії утворю ються вакуолі, які з часом зливаються у великі лакуни — лакунарна стадія розвитку трофобласта. Цим часом на протилежному полюсі бластоцисти плоскі клітини, мож ливо, гіпобластичного походження, формують тонку екзоцеломічну мембрану — мем брану Гейзера, яка вистеляє внутрішню поверхню цитотрофобласта. Мембрана Гей зера разом із гіпобластом утворює вистілку екзоцеломічної порожнини — первинного
жовткового мішка.
На 11–12 ту добу розвитку бластоциста повністю занурюється у строму ендомет рія, а поверхневий епітелій закриває первинний дефект ендометрія. Бластоциста дещо випинається у просвіт матки. Трофобласт утворює лакунарні простори в син цитії, які сполучаються між собою з утворенням сітки. Ця сітка найбільш виражена на ембріональному полюсі, тимчасом як на протилежному полюсі трофобласт утво рений переважно клітинами цитотрофобласта. Клітини синцитіотрофобласта про никають у строму ендометрія і руйнують едотеліальний шар материнських капі лярів. Ці капіляри стають розширеними, синусоїдними. Синцитіальні лакуни сполу чаються з синусоїдними капілярами, і материнська кров потрапляє у лакунарну сис тему трофобласта. При руйнуванні трофобластом все більшої кількості синусоїдів материнська кров починає циркулювати через лакунарну систему, виникає матко? во?плацентарний кровотік. Між внутрішньою поверхнею цитотрофобласта і зовніш ньою поверхнею екзоцеломічної порожнини з’являється нова популяція клітин. Ці клітини походять із жовткового мішка і утворюють пухку сполучну тканину — поза? зародкову мезодерму, яка заповнює всі простори між трофобластом зовні та амніо ном і екзоцеломічною мембраною зсередини. Незабаром у позазародковій мезодермі виникають великі порожнини, при злитті яких утворюється простір — позазародко вий целом, або порожнина хоріона. Цей простір оточує первинний жовтковий мішок і амніотичну порожнину, за винятком тієї ділянки, де зародковий диск прикріплюєть ся до трофобласта сполучною ніжкою. Та частина позазародкової мезодерми, що вистеляє цитотрофобласт і амніон, називається позазародковою соматоплевральною мезодермою. А частина, що вкриває жовтковий мішок, має назву позазародкової спланхноплевральної мезодерми. Ріст двошарового зародкового диска відбувається відносно повільно. Клітини ендометрія в цей час стають полігональними, накопичу ють глікоген і ліпіди, міжклітинні простори накопичують рідину, тканина стає на бряклою — відбувається децидуальна реакція, яка спочатку охоплює місце імплан тації, а потім поширюється на весь ендометрій.
До 13 ї доби дефект на поверхні ендометрія загоюється, але інколи з місця імплан тації може виникати кровотеча внаслідок збільшення кровотоку в лакунарних про сторах. Ця кровотеча відбувається близько 28 го дня циклу і може сприйматися як менструація, що призводить до неточного визначення гестаційного віку.
Клітини цитотрофобласта проліферують і вростають у синцитіотрофобласт, фор муючи оточені синцитієм клітинні колонки — первинні ворсинки. З гіпобласта тим часом виселяються клітини, які мігрують уздовж внутрішньої поверхні екзоцеломіч ної мембрани, проліферують і формують нову порожнину всередині екзоцеломічної порожнини — вторинний, або дефінітивний жовтковий мішок. Цей жовтковий мішок є значно меншим, ніж первинна екзоцеломічна порожнина, або первинний жовтко вий мішок. При формуванні дефінітивного жовткового мішка великі частини екзо целомічної порожнини стискаються і з них утворюються екзоцеломічні міхурці у позазародковому целомі — хоріонічні порожнини. Позазародкова мезодерма, що висте ляє всередині цитотрофобласт, отримує назву хоріонічної пластинки. Єдина ділянка,
155

Акушерство і гінекологія. Том 1
де позазародкова мезодерма пронизує хоріонічну порожнину, має назву сполучної ніжки, яка з розвитком кровоносних судин перетворюється на пупковий канатик.
У кінці другого тижня розвитку зародковий диск представлений двома прилег лими один до одного клітинними дисками: епібластом, який формує дно амніотич ної порожнини, що продовжує розширюватись, і гіпобластом, який формує дах вто ринного жовткового мішка. У головній ділянці гіпобласт утворює потовщення з циліндричних клітин, які щільно прилягають до диска епібласта — щічно глоткову перетинку.
Отже, другий тиждень розвитку відомий як тиждень двох: трофобласт диферен ціюється на два шари — цитотрофобласт і синцитіотрофобласт; ембріобласт утво рює два шари — епібласт і гіпобласт; позазародкова мезодерма розщеплюється на два шари — соматоплевру і спланхноплевру; формуються дві порожнини — амніо тична і жовткового мішка.
Клінічні кореляції. Імплантація є критичним періодом розвитку ембріона. Синци тіотрофобласт забезпечує секрецію гормонів, в тому числі хоріонічного гонадотропіну людини (ХГЛ), визначення якого в крові або сечі полягає в основі тестів на вагітність. Оскільки 50 % генів імплантованого ембріона мають батьківське походження, він є напівчужорідним для материнського організму. Але відторгнення зародка не відбу вається завдяки захисту комплексу факторів, які викликають продукцію цитокінів і протеїнів, а також специфічних молекул HLA–G, що належать до класу основного комплексу гістосумісності і блокують розпізнавання зародка як сторонньої тканини. За наявності у матері автоімунних захворювань (системний червоний вовчак, анти фосфоліпідний синдром) відторгнення ембріона відбувається за рахунок автоантитіл, які вторинно атакують зародок.
Аномальна імплантація може мати місце навіть у матці — не на передній або задній стінці, як звичайно, а біля внутрішнього зіва (передлежання плаценти). Імплантація поза маткою призводить до розвитку позаматкової, або ектопічної вагіт? ності — в матковій трубі (95 % випадків), в яєчнику, черевній порожнині.
Аномальні бластоцисти трапляються дуже часто. Більшість аномальних бласто цист не зумовлюють проявів вагітності, тому що їх трофобласт не здатний підтри мувати функцію жовтого тіла. Такі ембріони, ймовірно, відторгаються з черговою менструацією, і тому вагітність не настає. У деяких випадках аномальний розвиток трофобласта призводить до розвитку плацентарних оболонок за відсутності або за зменшення ембріональної тканини — і утворюється міхуровий занесок, один із варі антів гестаційної трофобластичної хвороби. Іншим варіантом цієї хвороби є розви ток злоякісної клітини з тканин трофобласта — хоріокарциноми. У разі утворення міхурового занеска його клітини є диплоїдними, але повністю мають батьківське походження, тобто має місце запліднення ооцита, в якому відсутнє ядро, з подаль шою дуплікацією чоловічих хромосом для відновлення диплоїдного набору. Ці дані також свідчать про те, що саме батьківські гени обумовлюють розвиток трофоблас та, оскільки при міхуровому занеску трофобласт формується навіть за відсутності жіночого пронуклеуса. Інші приклади функціональних відмінностей між материнсь кими і батьківськими генами дає вивчення генетичних хвороб, зумовлених похо дженням успадкованих (дефектного або втраченого) генів від батька чи від матері. Так, проявом успадкування делеції 15 ї хромосоми від батька є синдром Прадера — Віллі, тимчасом як той самий дефект, успадкований від матері, призводить до роз витку синдрому Ангельмана. Цей феномен диференційних модифікацій і експресії гомологічних алелів або хромосомних регіонів залежно від походження генетичного матеріалу відомий під назвою геномного імпринтингу. Імпринтинг стосується ауто сом і статевих хромосом (у всіх ссавців жіночої статі одна Х хромосома у соматич них клітинах є інактивованою) і модулюється шляхом метилювання ДНК. Такі за
156

Розділ 5. Репродуктивна генетика...
хворювання, як хорея Гентингтона, нейрофіброматоз, сімейні онкологічні захворю вання (пухлина Вільмса, сімейна ретинобластома) і міотонічна дистонія, а також синдром ламкої Х хромосоми також є проявами імпринтингу.
Репродуктивні втрати можуть відбуватись у пре і постімплантаційному періоді. Навіть у здорових фертильних жінок за оптимальних умов для настання вагітності 15 % ооцитів не запліднюються; 10–15 % починають дроблення, але не імпланту ються. Із 70–75 % зародків, що імплантувалися, лише 58 % доживають до другого тижня, з них 16 % є аномальними. Отже, до моменту затримки очікуваної менстру ації доживає лише 42 % яйцеклітин, що мали контакт зі сперматозоїдами. З них значна кількість ембріонів відторгається протягом наступних тижнів, і деяка кількість матиме аномалії при народженні.
Тришаровий зародковий диск. Гаструляція
Найхарактернішою подією третього тижня розвитку є гаструляція — утворення трьох зародкових листків — ектодерми, мезодерми і ендодерми ембріона (рис. 5.9). Гаструляція починається з утворення первинної смужки на поверхні епібласта. Го ловний кінець смужки — первинний вузлик — локалізується навколо первинної ямки. У ділянці первинного вузлика та первинної смужки клітини епібласта зміщуються досередини (інвагінують), утворюючи два нових клітинних шари — ендодерму і ме? зодерму. Клітини, які залишаються у складі епібласта, утворюють ектодерму. Отже, епібласт дає початок усім трьом зародковим листкам ембріона: ендодермі, мезодермі
іектодермі. Клітини внутрішньозародкової мезодерми мігрують доти, доки не зустрі нуться з позазародковою мезодермою, яка вкриває жовтковий мішок і амніон. У головному напрямку вони розходяться по обидва боки від прехордіальної пластин ки. Остання формується між верхівкою нотохондри та щічно глотковою мембраною
іє важливим фактором індукції розвитку переднього мозку.
Пренотохондріальні клітини інвагінують у первинну ямку, рухаючись у краніаль ному напрямку і досягають прехордіальної пластинки. Ці пренотохондріальні кліти ни вселяються в гіпобласт таким чином, що згодом серединна лінія ембріона склада ється з двох шарів клітин, які формують нотохордіальну пластинку. У міру заміщен ня гіпобласта клітинами ентодерми, які вростають усередину ембріона в ділянці первинної смужки, клітини нотохордіальної пластинки проліферують та відділя ються від ентодерми і згодом утворюють суцільний тяж клітин — дефінітивну ното? хорду, що лежить під нервовою трубкою і є зачатком осьового скелета. Спочатку формується краніальний кінець, потім первинна смужка поширюється у каудально му напрямку і утворює каудальний кінець ембріона. У місці, де ямка утворює загли бину в епібласті, утворюється нейрокишковий канал, що тимчасово сполучає порож нини амніона і жовткового мішка. На каудальному кінці ембріона з щільно зроще них клітин ектодерми і ентодерми, без проміжного шару мезодерми, утворюється клоакальна мембрана. Після появи клоакальної мембрани, близько 16 го дня розвит ку, задня ділянка стінки жовткового мішка формує невеликий дивертикул, який вростає у сполучну ніжку — алантокишковий дивертикул, або алантоїс. У людини алантоїс залишається рудиментарним і може спричинювати вади розвитку сечового міхура.
Перед початком і в процесі гаструляції відбувається становлення осей тіла: пере дньозадньої, дорзовентральної і лівоправосторонньої. Клітини задньої крайової зони (PMZ) на каудальному кінці ембріона виділяють активінподібну речовину сімей ства генів, трансформуючих фактори росту β (TGF β ), яка індукує утворення пер винної смужки. Цим визначається краніально каудальна вісь зародка. Після утво рення первинної смужки низка генів регулює формування дорзальної та вентраль
157

Акушерство і гінекологія. Том 1
Нотохорда |
Амніотична |
|
|
порожнина |
Приосьова |
Проміжна |
|
|
Ектодерма |
мезодерма |
мезодерма |
|
|
|
Міжклітинні |
|
Мезодерма |
простори |
|
|
Спинна |
у бічній |
|
|
пластинці |
||
|
|
аорта |
|
|
|
|
|
|
|
Проміжна |
|
Амніон Нервова борозна |
мезодерма |
Соміт |
|
Парієтальний
листок
мезодерми
Вісцеральний
листок
мезодерми
Внутрішньо зародкова порожнина тіла
Ендодерма
Рис. 5.9. Розвиток мезодермального зародкового листка (17 та–21 ша доба)
ної мезодерми, а також головних і хвостових структур. Інший ген сімейства TGF β , кістковий морфогенетичний протеїн 4 (ВМР 4), продукується на всій площі ембрі онального диска. У присутності цього протеїну та фактора росту фібробластів (FGF) мезодерма вентралізується, забезпечуючи утворення нирок (проміжна мезодерма), крові, а також мезодерми стінки тіла (мезодерма бічної пластинки). Хордин, ногін і фолістатин пригнічують активність ВМР 4 і забезпечують дорзалізацію мезодерми, внаслідок чого утворюються нотохорда і сомітомери ділянки голови. За відсутності фактора HNF 3β ембріони не здатні до нормальної гаструляції та не мають перед нього та середнього мозку. Ген Goosecoid активує інгібітори ВМР 4 і регулює розви ток голови. Надлишкова або недостатня експресія цього гена призводить до тяжких вад розвитку голови, включаючи її подвоєння.
Утворення дорзальної мезодерми в серединній і каудальній ділянках регіонах регулює ген Brachyury (Т). Відсутність цього гена спричинює вкорочення ембріо нальної осі — каудальний дисгенез. Ступінь вкорочення залежить від терміну ви никнення дефіциту цього протеїну.
Встановлення лівоправосторонньої асиметрії регулюється каскадом генів, у тому числі sonic hedgehog (Shh), продукт активності якого надходить до лівої частини первинного вузлика за допомогою рецептора активіну ІІа, експресію якого здійснює права частина первинного вузлика. Протеїн Shh активує гени nodal і lefty, які прояв ляються у мезодермі бічної пластинки лівої сторони зародка і регулюють активність інших генів для забезпечення асиметрії так, що серце і селезінка опиняються у лівій половині тіла, а більша частина печінки — у правій.
Клітини епібласта, які мігрують через первинний вузлик і смужку, предетерміно вані своїм місцем знаходження до утворення певних типів мезодерми і ендодерми. Так, клітини, що вростають через краніальну ділянку первинного вузлика, стають
158