

2. Морфология и формирование гематоенцефалічного барьера
Морфологическая локализация ГЕБ была уточнена и научно обоснована в конце 60-х годов. Классические опыты убедительно показали, что, как при внутривенном, так и при інтравентрикулярному введении экспериментальным животным ультраструктурних индикаторов - пероксидази хрону, миелопероксидазы, коллоидного лантанума - обмен веществ в обоих направлениях прекращается на уровне плотных межэндотелиальных соединений капилляров мозга, которые являются структурной основой ГЕБ.
В настоящее время теоретически и экспериментально обоснована морфологическая организация ГЕБ, которая включает несколько уровней клеточных систем (рис. 2.1). Первый уровень - двомембранний слой эндотелиоцитов, второй, - базальная мембрана, которая имеет фибриллы и клеточные компоненты (перицити), третий - астроцитарна муфта, которая укрывает 85-90% поверхности ГЕБ и образованная отростками астроцитов.
Строение ГЕБ с некоторыми вариациями сохраняется практически во всех отделах головного мозга, кроме гіпоталамо-гіпофізарної области, где базальная мембрана имеет перикапілярні пространства, а сам барьер есть значительно фенестрованим.
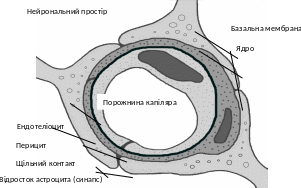
Рис 2.1. Схема поперечного среза капилляру ГЕБ.
В отличие от общей системы циркуляции крови, капилляры головного мозга характеризуются крепким межэндотелиальным соединением, отсутствием пор и фенестрів между эндотелиоцитами, сплошной базальной мембраной. Диффузия метаболітів и ионов в направлении кровь - мозг осуществляется на очень короткой дистанции. Расстояние между отдельными нейронами и капилляром крови составляет лишь несколько диаметров клетки. Кроме того, каждый сегмент сосуда часто образован единичной эндотелиальной клеткой. Эндотелиоциты, плотно контактируя друг с другом, образуют своеобразный капиллярный канал с очень тонкой стенкой (рис. 2.2). Базальная мембрана полностью укрывает эндотелиальные клетки. Базальный слой и эндотелий контактируют с пресинаптической мембраной отростков астроцитов. Диффузия метаболітів, лекарственных веществ между всей системой циркуляции крови и нейронами обеспечивается транспортными системами эндотелий - астроглія/міжклітинний пространство - нейрон. Отмечено, что нарушение проницаемости клеточной системы ГЕБ приводит к изменению нейронального метаболизма, функциональных расстройств, а в дальнейшем и к органическому поражению ЦНС.Щильни межэндотелиальные контакты неповрежденного ГЕБ ограничивают диффузию в мозг веществ больше 10 - 15 нм в диаметре.
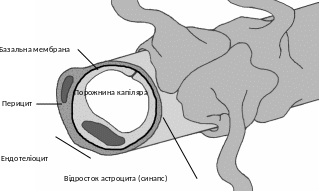
Рис. 2.2. Пространственная модель капилляра ГЕБ.
Прочность межэндотелиальных контактов обусловлена высокомолекулярными белками кадгеринами (Р-кадгерини), что опосредствуют Ca2+-залежну клеточную адгезию. Р-кадгерини преимущественно включены в структуру мембран эндотелиоцитов ГЕБ по всей длине межэндотелиальных соединений.
Следующий характерный признак эндотелия капилляров мозга - низкая степень трансцитозу (пиноцитозу). Это связано, по данным ряда авторов, с низким содержимым цитоплазматических везикул, например, по сравнению с м’язевими капиллярами, и отсутствием в эндотелии ГЕБ белка сіалоглікопротеїну dp60. Последний обеспечивает процесс пиноцитоза и присутствует в эндотелии сосудов всех других типов.
Мембраны эндотелия, которые контактируют с кровью, получили название люмінальні, а те, которые контактируют с мозговой тканью, - аблюмінальні (антилюмінальні) (от лат. lumen - просвет). Такое разделение связано с особенностями распределения разных за функциональным значением ферментных систем. При повреждении эндотелиальных клеток аблюмінальний барьер энзима, который хранит функциональную способность, продолжает выполнять защитные функции.
Перицити для ГЕБ является аналогами гладких мышц, поддерживая тонус базальной мембраны и выполняя сократительную функцию. Особенностью их функционирования считается экспрессия церебральными перицитами фермента аминопептидазы (AP-N), которая расщепляет N-кінцеву участок олигопептидов. Функциональный контакт перицитів одновременно с эндотелием и отростками астроцитов позволил допустить двоякую роль церебральной аминопептидазы: с одной стороны, участие в катаболизме пептидов, который ограничивает их доступ в направлении кровь - мозг, с другой стороны - обеспечения інактивації нейротрансмиттеров синаптосомальних зон и их экскрецию в направлении мозг - кровь. Одной из основных функций перицитів есть их влияние на регенерацию эндотелия ГЕБ, который опосредствует секрецией TGF ?.
Следующей функционально значимой единицей ГЕБ является астроглія. Экспериментальные работы подтвердили многофункциональность астроцитов, их участие в регуляции обмена нейротрансмиттеров, стимуляции синтеза миелина олігодендроцитами, в иммунном ответе мозга, активном транспорте ряда ионов. Кроме того, доказано непосредственное влияние астроглії на процессы дифференциации клеток, связанные с формированием эндотелия ГЕБ. Основной вопрос онтогенеза, который касается формирования ГЕБ, заключался в уточнении генетической переддетермінованості дифференциации эндотелиоцитов или наличия определенных эпигенетических факторов, которые индуктируют характерное развитие ГЕБ в генетически поліпотентному васкулярному эндотелии. Решению данного вопроса способствовали оригинальные опыты по трансплантации ткани мозга эмбриона перепела в целомическую полость куриного эмбриону. Тем самым была созданная модель мозга перепела, васкуляризированного сосудами кишечника цыпленка. Последующее морфологическое исследование новообразованных капилляров мозга перепела показало, что периферические эндотелиоциты приобрели фенотипових черты ГЕБ. Данный экспериментальный подход позволил сделать вывод, во-первых, о генетически поліпотентні возможности эндотелия, во-вторых, о способности мозговой ткани индуктировать специфические черты ГЕБ в гетерогенных васкулярно-ендотеліальних клетках.
Полученные результаты подтвердили вывод о генетической поліпотентність эндотелия и влияние на его дифференциацию микроокружения ткани мозга. В то же время описанные экспериментальные исследования не уточняли ни индуктивный тип клеток мозга, ни механизм взаимодействия разных клеточных систем ГЕБ.
Эксперименты из моделирования ГЕБ in vitro, выполненные в последние годы на высоком научно методическому уровни, убедительно продемонстрировали, что культура эндотелиоцитов, полученная из мозговых капилляров, хранит фенотип ГЕБ только при пассивировке последних с астроцитами. Только культуры эндотелиальных клеток с клетками астроглії, или обогащение питательной среды астроцитами, обусловливали и сохраняли специфические свойства ГЕБ, такие как прочность межэндотелиальных контактов, экспрессия эндотелиоцитами ?-глутамілтранспептидази (маркер только эндотелию ГЕБ), усвоение глюкозы, повышения в 3 раза активности липопротеиновых рецепторов мембран, стимулирования регенерации эндотелия. Кроме того, культура клеток мозга обогащена астроцитами обнаруживала более выраженный экранирующий эффект относительно токсичных веществ, храня целостность нейронов.
Позже была подтвержденная возможность экспрессии астроглією растворимого астроцитарного фактора, способного индуктировать специфические черты ГЕБ при взаимодействии в периферическом эндотелии. Именно астроглія благодаря секреции определенных растворимых факторов и множественности контактов является интегрирующим субстратом между разными типами клеток мозга. Таким образом, астроциты регулируют развитие и формирование ГЕБ, обеспечивают сохранение его фенотипу, способствуют регенерации его эндотелия при повреждении, является компонентами транспортной системы для метаболітів нейрональних структур.