

3). Экология круглых червей. Понятие о биогельминтах. Морфология, жизненный цикл, патогенное действие трихинеллы. Диагностика и профилактика трихинеллеза.
Тип: Круглые черви (Nemathelminthes)
Класс: Собственно круглые черви (Nematoda)
Вид Заболевание
Ascaris lumbricoides аскаридоз
(аскарида человеческая)
Toxocara canis (аскарида собачья) токсокароз
Enterobius vermicularis (острица) энтеробиоз
Trichocephalus trichiurus (власоглав) трихоцефалез
Trichinella spiralis (трихинелла) трихинеллез
Strongyloides stercoralis стронгилоидоз
(угрица кишечная)
Ancylostoma duodenale ( кривоголовка) анкилостомидоз
Necator americanus ( некатор) некатороз
Dracunculus medinensis (ришта) дракункулез
Dirofilaria repens (дирофилярия) дирофиляриоз
Filaria (филярии): филяриозы
Wuchereria bancrofti ( вухерерия) вухерериоз
Onchocerca volvulus ( онхоцерка) онхоцеркоз
Brugia malayi ( бругия) бругиоз
Согласно классификации В.А. Догеля (1981), тип Nemathelminthes включает в себя пять классов:
- 1 Класс Nematoda (Нематоды).
- 2 Класс Gastrotricha (Брюхоресничные).
- 3 Класс Gordicea (Волосатики).
- 4 Класс Kinorinchi (Киноринхи).
- 5 Класс Rotatoria (Коловратки).
Становление типа круглые черви связано с рядом ароморфозов и усложнением систем органов. Для них характерно:
- первичная полость тела, заполненная полостной жидкостью, которая выполняет роль гидроскелета и придает упругость и эластичность телу паразита;
- наличие в пищеварительной системе заднего отдела кишечника эктодермального происхождения, что обеспечивает непрерывное пищеварение и продвижение пищи в одном направлении;
-раздельнополость с выраженным половым диморфизмом. Самки крупнее самцов. Задний отдел тела самцов загнут на брюшную сторону.
В перечисленных классах только класс круглые черви представляет группу, к которой относятся как паразиты человека, так и свободно живущие виды.
Морфологические особенности нематод. Большинство нематод имеет круглое, удлиненное, веретеновиднос тело, передний и задний концы которого заострены. Размеры круглых червей варьируют в очень широких пределах, от микроскопически малых, не превышающих в длину 0,3 мм, до 6 - 8 м. (Placentonema gigantissima).
Ротовое отверстие располагается терминально на передней конце. Анальное отверстие локализуется субтерминально на вентральной поверхности. Женское половое отверстие лежит в передней половине тела вентрально. Самостоятельное мужское половое отверстие у всех видов круглых червей отсутствует.
Тело нематод покрыто кожно-мускульным мешком. Это сложное образование, состоящее из кутикулы, гиподермы, мускульных клеток. Кутикула представляет собой опорное скелетное образование. К ней прочно прикрепляется гладкая мускулатура. Под кутикулой залегает покровная ткань - гиподерма, состоящая из 4 утолщений: дорзального, вентрального и 2 латеральных. Гиподерма участвует в формировании кутикулы, имеет барьерную функцию. Под гиподермой находится продольная мускулатура.
Пищеварительная система. Хорошо развита, состоит из 3 отделов: переднего, среднего и заднего. Ротовое отверстие обычно окружено специальными придатками - губами, у более специализированных их число достигает 3-х. За пищеводом следует энтодермальная средняя кишка, далее - задний отдел кишечника. На границе между средним и задним отделами кишечника имеется сфинктер. Сквозная пищеварительная система заканчивается анальным отверстием.
Выделительная система. Представлена у нематод образованиями двух типов — шейными железами или реннетами, от которых идут длинные внутриклеточные каналы, они лежат в толще боковых гиподермальных валиков.
Нервная система круглых червей представлена нервным кольцом, состоящим из двух боковых ганглиев, соединенных комис-сурами. От кольца отходят продольные нервные стволы в количестве 8-12. Такая нервная система называется ортогональной.
Половая система круглых червей у разных представителей сильно варьирует. Женская половая система в подавляющем большинстве случаев парная. Она начинается двумя трубчатыми нитевидными яичниками, где формируются яйцеклетки. Яичники переходя в трубчатые яйцеводы с большим диаметром, затем следуют парные матки, за которыми следует непарный проток — влагалище. Мужская половая система представлена семенниками, где формируются сперматозоиды. За семенниками следует семяпровод, снабженный мускулистыми стенками. Семяизвергательный канал открывается в заднюю кишку, которая у самцов называется клоакой. Клоакн снабжена спикулами.
Кровеносная и дыхательная системы отсутствуют, что указывает на примитивность организации нематод. Дыхание совершается через покровы тела, либо биоэнергетический процесс протекает по типу аноксибиоза (брожения).
Происхождение круглых червей. Своеобразная организация круглых червей говорит о том, что они являются обособленной ветвью филогенетического древа животного мира, происходящей от одного из классов плоских червей - ресничных червей.
Биогельминты - это виды, личинки которых нуждаются в одном или нескольких промежуточных хозяевах. Например, трихинелла, ришта. Развитие круглых червей осуществляется с метаморфозом Переход от одной стадии к другой осуществляется в процессе четырех линек.
Трихинелла - Trichinella spiralis - возбудитель трихинеллеза. Трихинеллез встречается повсеместно, исключение составляет Австралия. Очаги заболевания сохраняются на Украине, в Белоруссии, Польше, США, ФРГ. В России групповые случаи заболевания зарегистрированы в Приморском, Красноярском краях, Воронежской, Челябинской областях. Это заболевание характеризуется природной очаговостью.
Морфологические особенности. Трихинелла имеет типичное для нематод строение. Самка трихинеллы имеет размеры 3 - 4 мм, ширина 0,06 мм, самец - 1,4 - 1,6 х 0,04 мм.
Тело паразита равномерно суживается кпереди. Отличитель ной особенностью является развитие одной половой трубки \ самки. Самец трихинеллы лишен спикул, их роль выполняют дна конусовидных кожистых выроста на заднем конце тела.
Цикл развития осуществляется с использованием дефинитивно го и промежуточного хозяина, которыми может быть одна особь, поражение этих хозяев осуществляется при поедании ими мяса живо: ных, содержащих инкапсулированные личинки трихинелл. Человек чаще всего заражается при употреблении в пищу свинины и мяса диких животных (кабанов, медведей, барсуков), не прошедших ветеринарного контроля. Инвазионные личинки в просвете кишечника освобождаются от окружающих их тканей и в течение трех суток достигают половой зрелости. Оплодотворенные самки прикренля ются головным концом тела к слизистой. Для трихинелл характерно яицеживорождение, так как из продуцируемых самкой яиц еще в матке вылупляются личинки, которые и выходят в просвет кишечника. Самка живет около 50 дней, отрождая 2 тыс. личинок. Самцы обычно погибают вскоре после копуляции. Личинки через стенку кишечника проникают в лимфатические сосуды, кровеносную систему, оказываясь в различных органах. Окончательно трихинеллы локализуются в поперечно-полосатой мускулатуре. Локализация личинок осуществляется в органах и тканях, особенно интенсивно снабжающихся кровью, кислородом и питательными веществами. Это мышцы языка, предплечья, диафрагмы, дельтовидные, межреберные, икроножные, мышцы глазного яблока, мочеиспускателыю! о канала, пищевода. На 3-й неделе личинки сворачиваются в спираль, вокруг которой в течение 2-3 месяцев формируется соединительнотканная капсула, стенки капсулы прорастают кровеносными сосу-_ дами и нервными окончаниями. С кровью паразит получает необходимые пищевые вещества и кислород, с кровью также удаляются и продукты обмена личинок. С 6 месяца наблюдается обызвествление капсул. Личинки при этом сохраняют жизнеспособность. Человек является биологическим тупиком в цикле развития трихинелл. Цир куляция паразита в окружающей среде осуществляется грызунами, хищными животными и птицами.
Патогенное действие. Трихинеллез начинается после инкубационного периода, ко торый длится от 7 дней до 5 недель. Начало заболевания острое. Развиваются характерные симптомы: отечность век или всего лип;: шеи, туловища, конечностей. Такая симптоматика обусловлена сил:, ной аллергической реакцией со стороны человека. Одновременно развивается лихорадка, достигающая максимума через 2-3 дня. Больного беспокоят боли в глазных, жевательных и шейных мышцах. Трихинеллез сопровождается расстройствами желудочно-кп шечного тракта, поносами, болями в животе, рвотой. Наблюдается бессонница, головные боли, депрессия или, наоборот, возбуждение, галлюцинации.
С образованием капсулы вокруг личинки больного беспокояi боли в пораженных мышцах (плечевых, поясничных, икроножных) может развиваться контрактура мышц.
Летальный исход при трихинеллезе связан с развитием осложнений в виде миокардита, менингоэнцефалита, пневмонии.' Прогноз при осложнениях неблагоприятен.
Диагностика основывается на биопсии мышц, где обнаруживаются личинки трихинелл. Большое значение имеет использование серологических реакций
Профилактические мероприятия сводятся к мерам личной и общественной профилактики.
Личная профилактика:
1. Не употреблять в пищу мясо домашних и диких животных, не прошедших ветеринарного обследования.
2. Покупать клейменное мясо, особенно свинину, на рынках, в магазинах.
Общественная профилактика:
1. Санитарно-просветительская работа, особенно в очагах.
2. Создавать санитарные условия, исключающие возможность заражения скота и других домашних животных.
3. Уничтожать крыс и мышей — основных источников трихинеллеза в еинантропных очагах.
4. Проведение должного санитарно-ветеринарного контроля (зараженные туши выбраковываются, уничтожаются или технически утилизируются).
4). Задача. Альбинизм наследуется как рецессивный аутосомныи признак. Заболевание встречается с частотой 1:20000. Вычислите количество гетерозигот в популяции.
Если принять, что альбинизм наследуется рессивно, то величина особей аа = q2 = 1/ 20000 . Отсюда частота аллеля а = q = 1/20 000 = 1/141. Тогда частота аллеля А = р = 1 – q = 1 – 1/141 = 140/141. Количество гетерозигот равно 2рq = 2 × 140/141 × 1/141 = 0,014, или 1,4 %.
Билет 23
1). Химические компоненты биологических систем. Роль неорганических соединений в нормальной жизнедеятельности клетки и организма. Металлы жизни. Последствия недостатка биогенных элементов для клеточного метаболизма.
Живые организмы и клетки – самовоспроизводящиеся химические системы,требующие связи разных групп, поэтому1. Основа клеточной химии – соединения углерода (способного соединятся в прочные цепи -С-С-Э, с легкими элементами, 99% занимают СНОNPS, 70% - вода), предмет органической химии, включая общие для всех 2- четыре основных типа молекул (с группами ОН,NН2, СН3 и С=О, Мг 100-1000):3. Сахара как пища для клеток, с формулой (СН2О)п- моно- и полисахариды, бывающие и скелетом и запасом энергии, при окислении(1У.Е)как и, из-за связи восстановления с гидрофобностью, (СН2)п-цепей, 4. Жирные кислоты - компоненты клеточных мембран, границ, в связи через остатки глицерина, с Ф- и +азотсодержащими, 5. Аминокислоты - субъединицы белков (сNН3+ и СОО- полюсами,источники разнообразных групп, зарядов и биоактивных веществ), 20 их кодируют(2х2)3также содержащие N+основание и Ф-6. Нуклеотиды - субъединицы ДНК и РНК, носитель и информации и энергии-полифосфатов, способные к автокатализу и воспроизводству, основы жизни. Заключение 2.2. Упорядоченность биологических систем и энергия 2.2.1. Упорядоченность биологических систем обусловлена выделением клеткой тепловой энергии, подобноТ-машинам, скорее холодильникам, создающим местный порядок и неравновесия за счет большего уравновешивания, окруженияисточников «более свободной»(см.2.4.1) энергии, первично – света 2- Солнца Фотосинтезирующими, для синтеза органических соединений (включая световые и темновые стадии,1 У.Е), их 3- Химическая энергия переходит от растений к животным, при обратной реакции питания-дыхания, 4. Клетки получают энергию в результате окисления биомолекул, 5- Распада, в результате последовательных ферментативных реакций (2.3, требуя катализа – уменьшения барьеров, энергии активации ферментами), запасая путем сопряжения (2.3.5, 2.4) 6- Часть энергии окисления в образовании АТР, «энерговалюте» (ЛИПМАН 1899-1953-86), а ее использование - 7. Гидролиз АТР обеспечивает упорядоченность в клетке (2.3, 2.4.2), в т.ч.синтез и транспорт молекул и движение клетки. Заключение 2.3. Питательные вещества и источники энергии клетки 1. Молекулы питательных веществ, расщепляясь в три этапа, образуют АТР (после гидролиза на мономеры и расщепления тех до С2-3-единиц, общих жирам, АК и углеводам), на этапе 2 - в процессе гликолиза (С6=2С3, брожения) образуемый и без кислорода(за счет ред-окс - большей выгоды СООН), но следующий этап 3. Окислительный катаболизм поставляет значительно больше энергии,- 4. Центральным процессом метаболизма является цикл лимонной кислоты
(КРЕБС Ханс, 1900-53-81, СЕНТ-ДЬЁРДЬИ 1893-1937-86)
- окисление ацетата (через соединение С-Н +С=О в С-С-ОН) до СО2 и Н-переносчиков, у эукариот – в митохондриях (гл.7), после 5.При окислительном фосфорилированииперенос электронов (Н) к кислороду приводит к образованию АТР и воды, посредством хемиосмотического сопряжения (ФОФ+НОН=Н+Ф-) с транспортом Н+, мембранным потенциалом(гл.7) 6. Аминокислоты и нуклеотиды принимают участие в круговороте азота (фиксируемого прокариотами и при высокой Т, молнии, и выводимого с мочевиной,аммиаком и др. Аналогично циклам С+-4иN-3/+5важны переходы S-2/+6, возможны и Г-1/+7) Заключение 2.4. Биосинтез и создание упорядоченности (транспорт внутрь и наружу связан с ката- и анаболизмом,разделением реакций и функций) 1.Возможность протекания реакции определяется величиной изменения свободнойэнергии (Гиббса или Гельмгольца, выражающей и энтропию, ее рост - 2-й закон ТД), возможны только уменьшающие ее, поэтому другие, как 2. Реакции биосинтеза (типа А-Н+В-ОН= АВ+НОН, обратно гидролизу, 5) часто сопряжены с гидролизом АТР (энергией отталкивания Ф-,аналогично отнятию воды Р2О5,с выделением ок.30 кДж/моль, для выгоды в сумме их энергий, общего изменения свободной, 1), фосфорилированием, аналогично 3. Коферменты участвуют в переносе специфических химических групп (из-за сложности получаясь нами извитаминов, см.НПХ,Б). Как переносчики ацетата (Ац-КоА) или Н (типа НАДН и ФАДН2, 2.3.4) в ОВ, ред-окс-реакциях, где при распаде образуются, а 4. Для биосинтеза необходимы восстановительные эквиваленты, чаще НАДФН. 5. Синтез био полимеров осуществляется путем повторения элементарных реакций дегидратации (за счет АТФ, фосфорилирования предшественников), обратно их гидролизу (2.3.1). Заключение 2.5. Координация катаболизма и биосинтеза- анаболизма, составляющих 1. Метаболизм - организуемый и регулируемый процесс, где тысячи реакций, в т.ч. конкурирующих, отражаемых картах («джунглях» безточных координат степени окисления-Сп) как 2- Метаболические пути регулируются изменениями ферментативной активности, при изменении условий, по принципу обратной связи (-ОС, например, конечным продуктом – первого фермента, при +ОС возможныавтокатализ иколебания), 3. Катаболические реакции могут обращаться при поглощении энергии (2.4.1), как распад и синтез глюкозы – гликолиз и глюконеогенез, регулируемые фосфофруктокиназой, активируемой АМФ, АДФ и ингибируемые – АТФ, ЖК и цитратом, обратно – бисфосфатазой. На большее время 4. Ферменты могут переходить в активное или неактивное состояния путем ковалентных модификаций, де-фосфорилирования – фосфатазами и протеинкиназами (ПК),
2). Полигенное наследование. Группы сцепления. Кроссинговер как механизм нарушения сцепления генов. Значение генетических рекомбинаций для человека.Тип наследования признаков, обусловленных действием многих генов, каждый из которых оказывает лишь слабое действие. Фенотипически проявление полигенно обусловленного признака зависит от условий внешней среды. У потомков наблюдается непрерывный ряд вариаций количественного проявления подобного признака, а не появление четко различающихся по фенотипу классов. В ряде случаев при блокировании отдельного гена признак не проявляется вообще, несмотря на его полигенную обусловленность. Это свидетельствует о пороговом проявлении признака.
Группы сцепления
Гены, находящиеся в одной хромосоме и наследующиеся сцепленно, составляют группу сцепления. Количество групп сцепления каждого вида должно соответствовать числу пар хромосом. Кроссинговер возникает со вполне определенной частотой для каждой пары генов, расположенных в одной группе сцепления. Причем, чем ближе в хромосоме расположены гены друг у другу, тем она выше. На основании анализа частоты кроссинговера между генами можно вычислить расстояние между генами и, таким образом, определить их локализацию в хромосоме План расположения генов в хромосоме называется картой хромосомы. Хромосомные карты построены для животных ряда видов, однако для собаки их нет, так как для их составления необходимы фундаментальные исследования генетики вида и большой статистический материал, получение которого у собак затруднительно. Для кошки установлено положение на хромосомах приблизительно для 50 генов (Бородин, 1995). На основании анализа результатов многочисленных экспериментов с мухой дрозофилой Т. Морган сформулировал свою хромосомную теорию наследственности, сущность которой заключается в следующем: 1. Материальные носители наследственности — гены — находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга. 2. Гены, расположенные в одной хромосоме, наследуются сцепленно. Число групп сцепления соответствует гаплоидному числу хромосом. 3. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно. 4. В потомстве гетерозиготных родителей новое сочетание генов, расположенных в одной паре хромосом, может возникнуть в результате кроссинговера в процессе мейоза. 5. Частота кроссинговера, определяемая по проценту кроссоверных особей, зависит от расстояния между генами. 6. На основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом. Сцепление генов приводит к одновременному наследованию блока признаков. Эти блоки могут сохраняться на протяжении нескольких поколений. Именно они способствуют передаче сложных признаков, например определяют внешнее сходство дедов и внуков, или обусловливают то, что все метисы колли или кокеров имеют совершенно определенную внешность независимо от внешнего вида второго родителя. Сцепление признаков может происходить в силу целого ряда причин, среди них и сцепление генов и плейотропия и действие определенных каналов морфогенеза. Сцепление признаков в значительной степени обусловливают тип собаки, препотентность производителя и делают возможным существование заводских линий.
Кроссинговер (от англ. crossing–over – перекрёст) – это обмен гомологичными участками гомологичных хромосом (хроматид).
Само явление перекрёста хромосом открыл Ф. Янссенс (1909, Бельгия), обнаруживший хиазмы в профазе I мейоза у саламандры. Однако теоретическая возможность кроссинговера была предсказана раньше.
Во-первых, американский цитолог У. Сэттон (1903) предположил, что в одной хромосоме может находиться несколько генов. В этом случае должно наблюдаться сцепленное наследование признаков, т.е. несколько разных признаков могут наследоваться так, как будто они контролируются одним геном. Тогда совокупность генов в одной хромосоме образует группу сцепления.
Во-вторых, в 1906 г. У. Бэтсон и Р. Пеннет обнаружили сцепленное наследование у душистого горошка. Они изучали совместное наследование: окраски цветков (пурпурная или красная) и формы пыльцевых зерен (удлиненная или округлая). При скрещивании дигетерозигот в их потомстве наблюдалось расщепление 11,1:0,9:0,9:3,1 вместо ожидаемого 9:3:3:1.
Таким образом, два признака (окраска цветков и морфология пыльцы) оказываются сцепленными. Соответственно, сцеплены и гены, контролирующие эти признаки («эффект взаимного притяжения»). Однако это сцепление не абсолютно: гены, контролирующие сцепленные признаки, могут приобретать независимость, и тогда появляются новые комбинации признаков. Вскоре (1911) были получены данные по сцепленному наследованию и нарушению сцепления некоторых признаков у дрозофилы.
Т. Морган (1910) и его ученик К. Бриджес (1916) доказали, что число групп сцепления соответствует гаплоидному числу хромосом. Опираясь на работы Янссенса и результаты гибридологического анализа, они связали явление перекрёста хромосом с феноменом нарушения сцепления генов. При перекрёсте хромосом происходит обмен между ними генетическим материалом (аллелями), и тогда происходит рекомбинация – появление новых сочетаний аллелей, например, AB + ab → Ab + aB.
Таким образом, термин «кроссинговер» употребляется и в цитологическом, и в генетическом значении.
В дальнейшем связь между перекрестом хромосом и появлением новых сочетаний признаков была подтверждена работами К. Дарлингтона, К. Штерна, Б. Мак-Клинток и др.
Цитогенетика кроссинговера
Механизм кроссинговера «разрыв–воссоединение»
Согласно теории Янссенса–Дарлингтона, кроссинговер происходит в профазе мейоза. Гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Ab и аВ. В анафазе первого деления мейоза происходит расхождение двухроматидных хромосом, а во втором делении – расхождение хроматид (однохроматидных хромосом). Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи. Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи.
Таким образом, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний (гаплотипов) наследственных задатков в хромосомах.
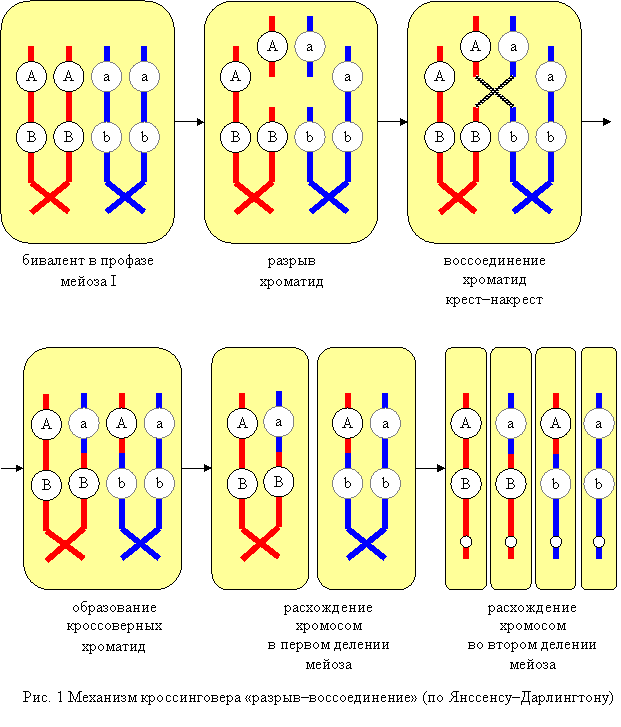
Примечание. Согласно другим теориям, кроссинговер связан с репликацией ДНК: или в пахитене мейоза, или в интерфазе (см. ниже). В частности, возможна смена матрицы в вилке репликации.
Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке происшедшего обмена. Рассмотрим пример, описанный в одной из ранних работ Моргана. Он исследовал частоту кроссинговера между генами w (white – белые глаза), у (yellow – желтое тело) и m (miniature – маленькие крылья), локализованными в Х-хромосоме D. melanogaster. Расстояние между генами w и у в процентах кроссинговера составило 1,3, а между генами у и m – 32,6. Если два акта кроссинговера наблюдаются случайно, то ожидаемая частота двойного кроссинговера должна быть равна произведению частот кроссинговера между генами уи w и генами w и m. Другими словами, частота двойных кроссинговеров будет 0,43%. В действительности в опыте был обнаружен лишь один двойной кроссинговер на 2205 мух, т. е. 0,045%. Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления фактически наблюдаемой частоты двойного кроссинговера на теоретически ожидаемую (при отсутствии интерференции) частоту. Он назвал этот показатель коэффициентом коинциденции, т. е. совпадения. Меллер показал, что в Х-хромосоме дрозофилы интерференция особенно велика на небольших расстояниях; с увеличением интервала между генами интенсивность ее уменьшается и на расстоянии около 40 морганид и более коэффициент коинциденции достигает 1 (максимального своего значения).
Двойной и множественный кроссинговер
Т. Морган предположил, что кроссинговер между двумя генами может происходить не только в одной, но и в двух и даже большем числе точек. Четное число перекрестов между двумя генами, в конечном счете, не приводит к их перемещению из одной гомологичной хромосомы в другую, поэтому число кроссинговеров и, следовательно, расстояние между этими генами, определенное в эксперименте, снижаются. Обычно это относится к достаточно далеко расположенным друг от друга генам. Естественно, что вероятность двойного перекреста всегда меньше вероятности одинарного. В принципе она будет равна произведению вероятности двух единичных актов рекомбинации. Например, если одиночный перекрест будет происходить с частотой 0,2, то двойной – с частотой 0,2 × 0,2 = 0,04. В дальнейшем, наряду с двойным кроссинговером, было открыто и явление множественного кроссинговера: гомологичные хроматиды могут обмениваться участками в трех, четырех и более точках.
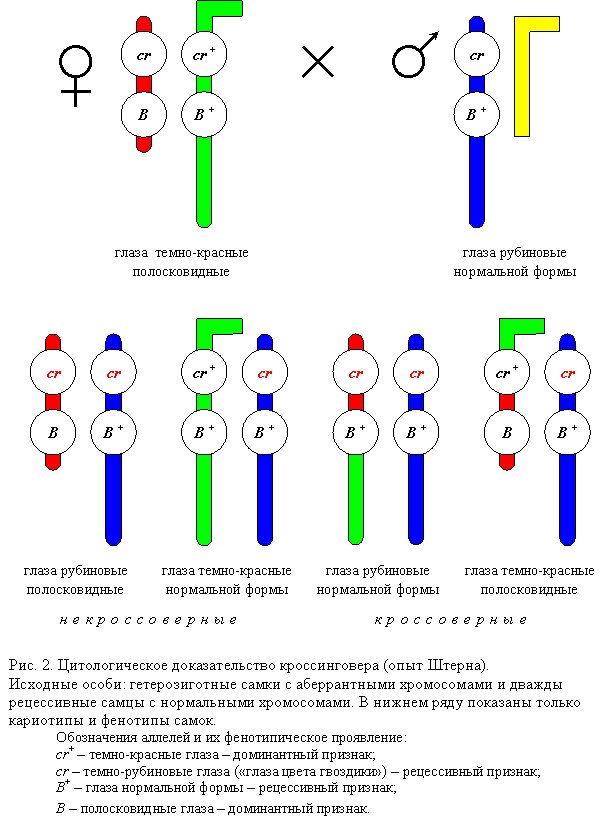
Цитологическое (цитогенетическое) доказательство кроссинговера
Прямые цитологические свидетельства обмена частей хромосом во время кроссинговера были получены в начале 30-х годов у дрозофилы и кукурузы. Рассмотрим опыт Штерна, проведенный на D. melanogaster. Обычно две гомологичные хромосомы морфологически неразличимы. Штерн исследовал Х-хромосомы, которые имели морфологические различия и, следовательно, были гомологичны не полностью. Однако гомология между этими хромосомами сохранялась на большей части их длины, что позволяло им нормально спариваться и сегрегировать в мейозе (то есть нормально распределяться по дочерним клеткам). Одна из Х-хромосом самки в результате транслокации, т. е. перемещения фрагмента Y-хромосомы, приобрела Г-образную форму. Вторая Х-хромосома была короче нормальной, так как часть ее была перенесена на IV хромосому. Были получены самки, гетерозиготные по указанным двум, морфологически различным, Х-хромосомам, а также гетерозиготные по двум генам, локализованным в Х-хромосоме: Bar (В) и carnation (cr). Ген Bar – это полудоминантный ген, влияющий на количество фасеток и, следовательно, форму глаза (мутанты с аллелем В имеют полосковидные глаза). Ген cr контролирует окраску глаз (аллель cr+ обусловливает нормальную окраску глаз, а аллель cr – окраску глаз цвета красной гвоздики). Г-образная Х-хромосома несла аллели дикого типа В+ и cr+, укороченная хромосома – мутантные аллели В и cr. Самки указанного генотипа скрещивались с самцами, имевшими морфологически нормальную Х-хромосому с аллелями cr и В+. В потомстве самок было два класса мух с некроссоверными хромосомами (crB / crB+ и cr+B+ / crB+) и два класса мух, фенотип которых соответствовал кроссоверам (crB+ / crB+ и cr+B / crB+). Цитологическое исследование показало, что у кроссоверных особей произошел обмен участками Х-хромосом, и, соответственно, изменилась их форма. Все четыре класса самок имели по одной нормальной, т. е. палочковидной, хромосоме, полученной от отца. Кроссоверные самки содержали в своем кариотипе преобразованные в результате кроссинговера Х-хромосомы – длинную палочковидную или двуплечую с короткими плечами. Эти опыты, так же как и одновременно полученные аналогичные результаты на кукурузе, подтвердили гипотезу Моргана и его сотрудников о том, что кроссинговер представляет собой обмен участками гомологичных хромосом и что гены действительно локализованы в хромосомах.
Соматический (митотический) кроссинговер
В соматических клетках иногда происходят обмены между хроматидами гомологичных хромосом, в результате которых наблюдается комбинативная изменчивость, подобная той, которая регулярно генерируется мейозом. Нередко, особенно у дрозофилы и низших эукариот, гомологичные хромосомы синаптируют в митозе. Одна из аутосомно-рецессивных мутаций человека, в гомозиготном состоянии приводящая к тяжелому заболеванию, известному под названием синдром Блюма, сопровождается цитологической картиной, напоминающей синапсис гомологов и даже образование хиазм. Доказательство митотического кроссинговера было получено на дрозофиле при анализе изменчивости признаков, определяемых генами у (yellow – желтое тело) и sn (singed – опаленные щетинки), которые находятся в Х-хромосоме. Самка с генотипом ysn+ / y+sn гетерозиготна по генам у и sn, и поэтому в отсутствие митотического кроссинговера ее фенотип будет нормальным. Однако если кроссинговер произошел на стадии четырех хроматид между хроматидами разных гомологов (но не между сестринскими хроматидами), причем место обмена находится между геном sn и центромерой, то образуются клетки с генотипами y sn+ / y+ sn+ и y+sn/y+n. В этом случае на сером теле мухи с нормальными щетинками появятся близнецовые мозаичные пятна, одно из которых будет желтого цвета с нормальными щетинками, а другое – серого цвета с опаленными щетинками. Для этого необходимо, чтобы после кроссинговера обе хромосомы (бывшие хроматиды каждого из гомологов) y+ sn отошли к одному полюсу клетки, а хромосомы y sn+ – к другому. Потомки дочерних клеток, размножившись на стадии куколки, и приведут к появлению мозаичных пятен. Таким образом, мозаичные пятна образуются тогда, когда рядом расположены две группы (точнее, два клона) клеток, фенотипически отличающиеся друг от друга и от клеток остальных тканей данной особи.
Неравный кроссинговерЭто явление было детально изучено на примере гена Bar (В – полосковидные глаза), локализованного в Х-хромосоме D. melanogaster. Неравный кроссинговер связан с дупликацией какого-либо участка в одном из гомологов и с утратой его в другом гомологе. Обнаружено, что ген В может присутствовать в виде тандемных, т. е. следующих друг за другом, повторов, состоящих из двух и даже трех копий. Цитологический анализ подтвердил предположение о том, что неравный кроссинговер может вести к тандемным дупликациям. В области, соответствующей локализации гена В, на препаратах политенных хромосом отмечено увеличение числа дисков, пропорциональное дозе гена. Предполагается, что в эволюции неравный кроссинговер стимулирует создание тандемных дупликаций различных последовательностей и использование их в качестве сырого генетического материала для формирования новых генов и новых регуляционных систем.
Регуляция кроссинговераКроссинговер – это сложный физиолого-биохимический процесс, который находится под генетическим контролем клетки и подвержен влиянию факторов внешней среды. Поэтому в реальном эксперименте о частоте кроссинговера можно говорить, имея в виду все те условия, в которых она была определена. Кроссинговер практически отсутствует между гетероморфными Х- и Y-хромосомами. Если бы он происходил, то хромосомный механизм определения пола постоянно разрушался бы. Блокирование кроссинговера между этими хромосомами связано не только с различием в их величине (оно наблюдается не всегда), но и обусловлено Y-специфичными нуклеотидными последовательностями. Обязательное условие синапса хромосом (или их участков) – гомология нуклеотидных последовательностей.
Для абсолютного большинства высших эукариот характерна примерно одинаковая частота кроссинговера как у гомогаметного, так и гетерогаметного полов. Однако есть виды, у которых Кроссинговер отсутствует у особей гетерогаметного пола, в то время как у особей гомогаметного пола он протекает нормально. Такая ситуация наблюдается у гетерогаметных самцов дрозофилы и самок шелкопряда. Существенно, что частота митотического кроссинговера у этих видов у самцов и самок практически одинакова, что указывает на различные элементы контроля отдельных этапов генетической рекомбинации в половых и соматических клетках. В гетерохроматических районах, в частности прицентромерных, частота кроссинговера снижена, и поэтому истинное расстояние между генами в этих участках может быть изменено.
Обнаружены гены, выполняющие функции запирателей кроссинговера, но есть также гены, повышающие его частоту. Они иногда могут индуцировать заметное число кроссоверов у самцов дрозофилы. В качестве запирателей кроссинговера могут выступать также хромосомные перестройки, в частности инверсии. Они нарушают нормальную конъюгацию хромосом в зиготене.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток. Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
Биологическое значение кроссинговера Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Это означает, что…
а) в ходе естественного отбора в одних хромосомах происходит накопление «полезных» аллелей (и носители таких хромосом получают преимущество в борьбе за существование), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбывают из игры – элиминируются из популяций)
б) в ходе искусственного отбора в одних хромосомах накапливаются аллели хозяйственно-ценных признаков (и носители таких хромосом сохраняются селекционером), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбраковываются).
Эволюционное значение сцепленного наследования
В результате сцепления одна хромосома может содержать как благоприятные аллели (например, А), так и нейтральные или относительно неблагоприятные (например, N). Если некоторый гаплотип (например, AN) повышает приспособленность его носителей за счет наличия благоприятных аллелей A, то в популяции будут накапливаться как благоприятные аллели, так и сцепленные с ними нейтральные или относительно неблагоприятные N.
Пример. Гаплотип AN обладает преимуществом перед гаплотипом «дикого типа» (++) за счет наличия благоприятного аллеля А, и тогда аллель N будет накапливаться в популяции, если он селективно нейтральный или даже относительно неблагоприятный (но его отрицательное влияние на приспособленность компенсируется положительным влиянием аллеля А).
Эволюционное значение кроссинговера
В результате кроссинговера неблагоприятные аллели, первоначально сцепленные с благоприятными, могут переходить в другую хромосому. Тогда возникают новые гаплотипы, не содержащие неблагоприятных аллелей, и эти неблагоприятные аллели элиминируются из популяции.
Пример. Гаплотип Al оказывается неблагоприятным по сравнению с гаплотипом «дикого типа» (++) за счет наличия летального аллеля l. Поэтому аллель А(благоприятный, нейтральный ил несколько снижающий приспособленность) не может проявиться в фенотипе, поскольку данный гаплотип (Al) содержит летальный аллель l. В результате кроссинговера возникают рекомбинантные гаплотипы A+ и +l. Гаплотип +l элиминируется из популяции, а гаплотип A+ фиксируется (даже в том случае, если аллель А несколько снижает приспособленность его носителей).