

8.12. Патология белкового обмена
В предыдущих разделах изложены нарушения обмена серосодержащих аминокислот (обмен серосодержащих аминокислот), нарушения обмена фенилаланина, тирозина (обмен фенилаланина и тирозина), нарушения синтеза мочевины (биосинтез мочевины).
Белковое голоданиеможет наблюдаться при недостатке белков в рационе, недостатке витаминов, заболеваниях желудочно–кишечного тракта. При белковом голодании усиливается распад собственных тканевых белков, особенно в таких тканях как мышцы, печень и плазма крови. Развивается отрицательный азотистый баланс, дистрофия мышц, нарушение антитоксической функции печени, гипопротеинемия и как следствие, «голодные» отёки. Происходит уменьшение подкожной клетчатки, страдают иммунные процессы. Тяжелая форма белкового голодания носит названиеквашиоркор(красный ребёнок).
9. Обмен и функции нуклеиновых кислот
9.1. Химический состав и строение нуклеиновых кислот
Нуклеиновые кислоты – высокомолекулярные полимерные соединения, мономером которых являются мононуклеотиды.
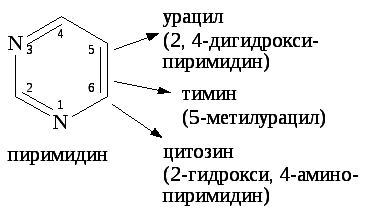
Мононуклеотиды включают в свой состав три вещества: пуриновые (аденин, гуанин) и пиримидиновые (урацил, тимин, цитозин) азотистые основания, углевод (рибоза или дезоксирибоза), фосфорную кислоту.
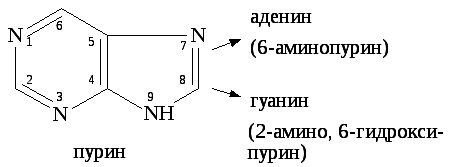
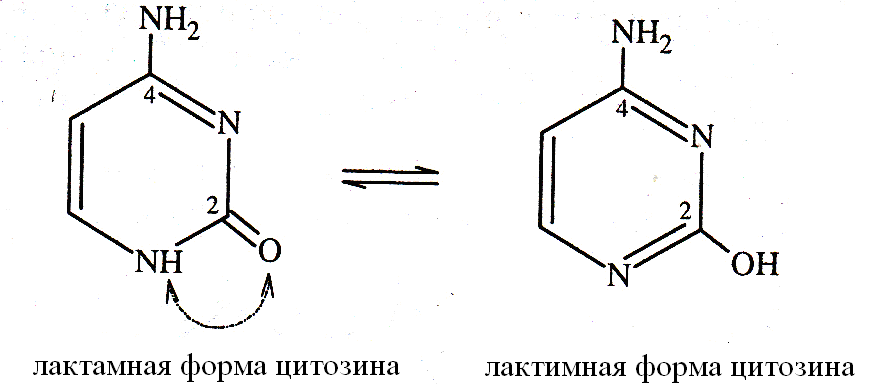
Гуанин, содержащий в шестом положении кольца ОН группу, всоставе нуклеиновых кислот находится в лактамной форме (=О). При образовании лактамной формы атом водорода перемещается от кислорода к рядом расположенному первому атому азота.

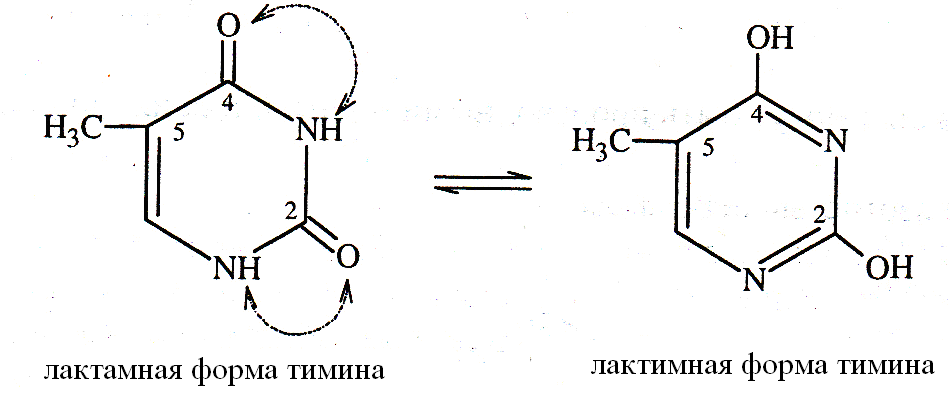
Пиримидиновые нуклеотиды в составе нуклеиновых кислот находятся в лактамной форме, позволяющей пиримидиновым основаниям включаться в состав нуклеотидов и участвовать в образовании водородных связей в ДНК.
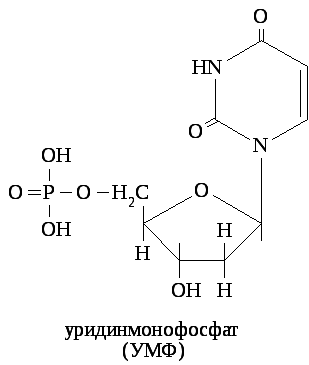
Азотистые основания соединяются с углеводом β - N- гликозидной связью. Пуриновое основание соединяется с углеводом за счёт 9 атома азота, а пиримидиновое - за счёт 1 атома азота. К углеводу фосфоэфирной связью присоединяется фосфорная кислота.
В зависимости от количества остатков фосфорной кислоты различают нуклеозидмонофосфаты (цикло-АМФ, ФАФС, S-аденозилметионин), нуклеозиддифосфаты (УДФ-глюкоза, ЦДФ-холин), нуклеозидтрифосфаты (макроэрги).

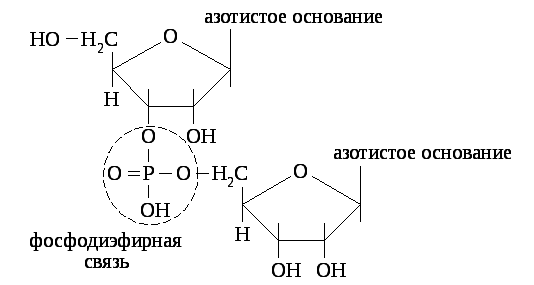
Нуклеотиды соединяются в полинуклеотидную цепь фосфодиэфирными связями, образованными фосфорной кислотой и рибозой соседних нуклеотидов (к рибозе одного нуклеотида фосфорная кислота присоединяется в третьем положении, а к рибозе соседнего нуклеотида - в пятом положении).
Рибонуклеиновые кислоты (РНК)
Все виды РНК состоят из одной полинуклеотидной цепи. Азотистые основания в РНК представлены аденином, гуанином, урацилом, цитозином. Углевод представлен рибозой. Различают несколько видов РНК.
Т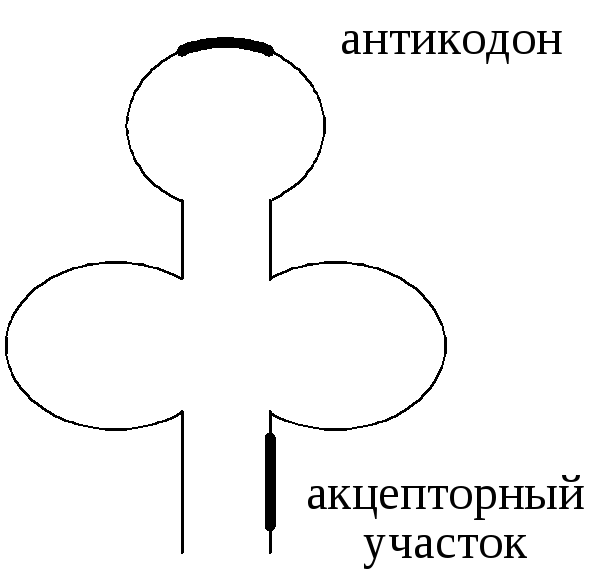 ранспортные
РНК (тРНК) осуществляют транспорт
аминокислот на рибосомы, к месту синтеза
белка. Молекулярная масса тРНК составляет
около 30 000 д. Полинуклеотидная цепь тРНК
включает в свой состав 70-80 нуклеотидов.
На долю тРНК приходится около 15% всего
запаса РНК клетки. В тРНК содержится
большое количество минорных нуклеотидов
(модифицированные формы обычных
оснований). Между комплементарными
нуклеотидами участков тРНК возникают
водородные связи, и формируется её
вторичная структура в виде трилистника
(форма «клеверного листа»). В составе
тРНК выделяют 2 важных функциональных
участка. На средней петле располагается
антикодон, комплементарный кодонам
иРНК, на открытом конце – акцепторный
участок, к которому присоединяется
аминокислота. Для переноса каждой
аминокислоты в клетках имеется «своя»
тРНК.
ранспортные
РНК (тРНК) осуществляют транспорт
аминокислот на рибосомы, к месту синтеза
белка. Молекулярная масса тРНК составляет
около 30 000 д. Полинуклеотидная цепь тРНК
включает в свой состав 70-80 нуклеотидов.
На долю тРНК приходится около 15% всего
запаса РНК клетки. В тРНК содержится
большое количество минорных нуклеотидов
(модифицированные формы обычных
оснований). Между комплементарными
нуклеотидами участков тРНК возникают
водородные связи, и формируется её
вторичная структура в виде трилистника
(форма «клеверного листа»). В составе
тРНК выделяют 2 важных функциональных
участка. На средней петле располагается
антикодон, комплементарный кодонам
иРНК, на открытом конце – акцепторный
участок, к которому присоединяется
аминокислота. Для переноса каждой
аминокислоты в клетках имеется «своя»
тРНК.
тРНК имеет 3-ю структуру, которая представляет собой компактное наложение петель друг на друга.
Различают изоакацепторные тРНК, которые переносят одну и ту же аминокислоту, но отличаются структурой и одним нуклеотидом в антикодоне.
Рибосомальные РНК (рРНК)составляют до 80% всей РНК клетки. Рибосомальная РНК локализована в рибосомах и обеспечивает биосинтез белка. Рибосома человека имеет молекулярную массу 80S, включает большую субъединицу массой 60S(включает РНК с массой 5S, 5,8S, 25S) и малую субъединицу с массой 40S(включает РНК массой 18S). Вторичная структура рРНК представляет компактную укладку, формирующую овальный каркас большой и малой субъединиц, соединённый с белками.
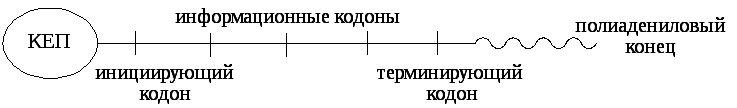
Информационная РНК (иРНК)имеет большую молекулярную массу около 106д. На её долю приходится около 3% РНК клетки. В первичной структуре иРНК представлены кодоны, среди которых различают инициирующие (в них первый нуклеотид А), терминирующие (в них первый нуклеотид У) и кодоны, определяющие порядок аминокислот в синтезируемом белке. На одном конце иРНК имеется «шапочка - кеп», необходимая для связывания иРНК с малой субъединицей рибосомы в начале синтеза белка. На другом конце полинуклеотидной цепи располагается полиадениловый «хвост», выполняющий защитную функцию.
Дезоксирибонуклеиновая кислота
Азотистые основания в ДНК представлены аденином, гуанином, тимином, цитозином, углевод - дезоксирибозой. ДНК играет важную роль в хранении генетической информации. В отличие от РНК в ДНК присутствуют две полинуклеотидные цепи. Молекулярная масса ДНК около 109 д. Полинуклеотидные цепи в молекуле ДНК объединяются по принципу комплементарности, известному как «правила Чаргафа» (1939 г.). Правила Э. Чаргафа включают несколько положений.
Количество остатков аденина равно количеству остатков тимина (А=Т). Количество остатков гуанина равно количеству остатков цитозина (Г=Ц).
Сумма пуриновых оснований (А + Г) равна сумме пиримидиновых оснований (Т + Ц).
В комплементарных позициях количество оснований с аминогруппой равно количеству оснований с кетогруппой (Г + Т = А + Ц).
Для всех видов ДНК существует коэффициент видовой специфичности – отношение Г + Ц/А + Т < 1.
На основании данных правил и результатов рентгеноструктурного анализа Д. Уотсон и Ф. Крик создали модель структуры ДНК, согласно которой в молекуле ДНК две полинуклеотидные цепи располагаются антипараллельно и соединяются водородными связями между комплементарными азотистыми основаниями. В паре А-Т возникают 2 водородные связи, в паре Г - Ц формируются 3 водородные связи.
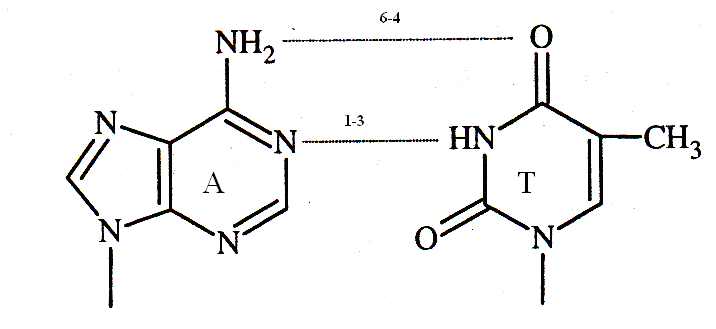
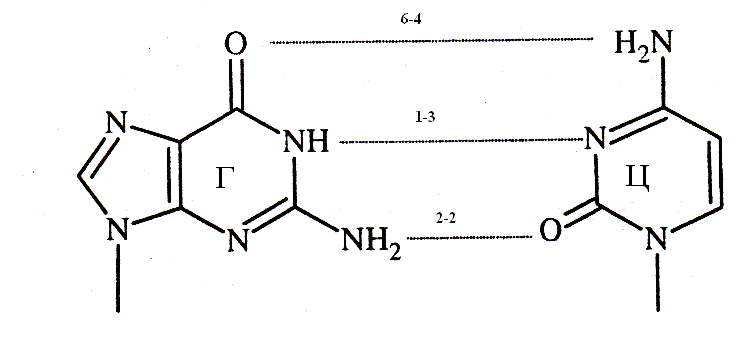
Вторичная структура наиболее распространённой В - формы ДНК представляет собой правозакрученную двойную спираль, в одном витке которой уложено 10 пар нуклеотидов. Шаг спирали равен 3,4 нм. В фагах присутствует Zформа (зигзагообразная) и кольцевая форма ДНК.
Для ДНК характерна более компактная укладка в виде суперспирали. ДНК формирует надмолекулярную структуру, объединяясь с ядерными белками и формируя полинуклеосомы.
ДНК образует вязкие водные растворы с двойным лучепреломлением, поглощающие УФЛ в интервале 260-280 нм. Под действием концентрированных растворов кислот, мочевины, физических факторов возможна денатурация ДНК, при которой происходит разрыв водородных связей и расхождение цепей молекулы ДНК. Денатурация может носить обратимый характер и переходить в ренативацию – восстановление водородных связей и структуры ДНК после удаления денатурирующего фактора. При ренативации возможно явление гибридизации – объединение комплементарных цепей ДНК из разных организмов. Возможна гибридизация нитей ДНК с РНК.