10.3. Механизмы действия гормонов
Различают несколько механизмов реализации гормонального сигнала для водорастворимых и водонерастворимых гормонов.
Все гормоны оказывают три конечных эффекта:
изменение количества белков и ферментов за счёт изменения скорости их синтеза;
изменение активности имеющихся в клетки ферментов;
изменение проницаемости клеточных мембран.
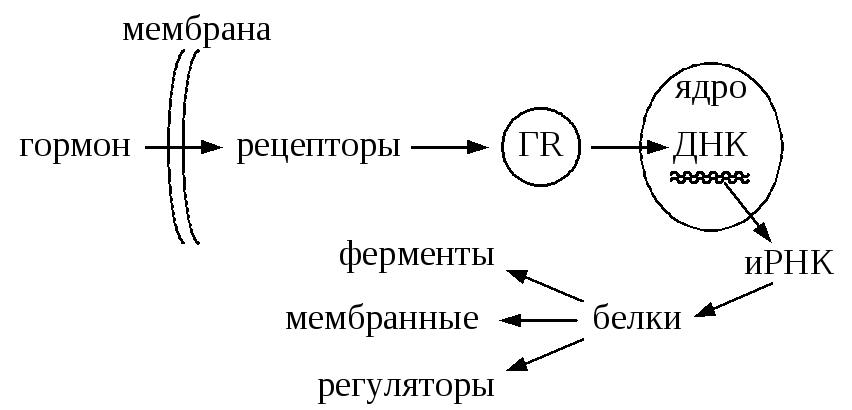
10.3.1. Цитозольный механизм действия гидрофобных (липофильных) гормоновЛипофильные гормоны способны проникать в клетку через клеточную мембрану, поэтому рецепторы для них располагаются внутриклеточно в цитозоле, на митохондриях, на поверхности ядра. Рецепторы гормонов чаще всего включают 2 домена: для связывания с гормоном и для связывания с ДНК. Рецептор при взаимодействии с гормоном изменяет свою структуру, освобождается от шаперонов, в результате чего гормон - рецепторный комплекс приобретает способность проникать внутрь ядра и взаимодействовать с определёнными участками ДНК. Это, в свою очередь, ведёт к изменению скорости транскрипции (синтез РНК), а вследствие этого меняется и скорость трансляции (синтез белка).
10.3.2. Мембранный механизм действия водорастворимых гормонов
Водорастворимые гормоны не способны проникать через цитоплазматическую мембрану. Рецепторы для данной группы гормонов располагаются на поверхности клеточной мембраны. Поскольку гормоны не проходят внутрь клеток, между ними и внутриклеточными процессами необходим вторичный посредник, который передаёт гормональный сигнал внутрь клетки. В качестве вторичных посредников могут служить инозитолсодержащие фосфолипиды, ионы кальция, циклические нуклеотиды.
10.3.2.1. Циклические нуклеотиды - цАмф, цГмф - вторичные посредники
Гормон взаимодействует с рецептором и образует гормон - рецепторный комплекс, в котором меняется конформация рецептора. Это, в свою очередь, изменяет конформацию мембранного ГТФ - зависимого белка (G-белка) и ведёт к активации мембранного фермента аденилатциклазы, который переводит АТФ в цАМФ.
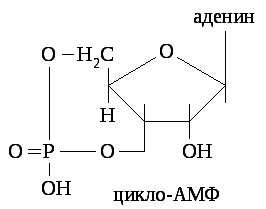
Внутриклеточный циклический АМФ служит вторичным посредником. Он активирует внутриклеточные ферменты протеинкиназы, которые катализируют фосфорилирование различных внутриклеточных белков (ферментов, мембранных белков), что приводит к реализации конечного эффекта гормона. Эффект гормона «выключается» под действием фермента фосфодиэстеразы, разрушающей цАМФ, и ферментов фосфатаз, дефосфорилирующих белки.
.

10.3.2.2. Ионы кальция - вторичные посредники
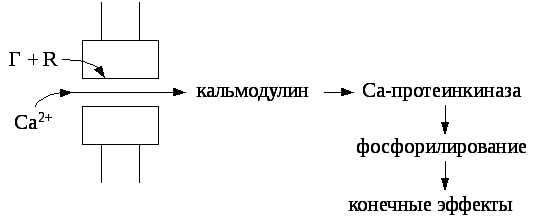
Взаимодействие гормона с рецептором повышает проницаемость кальциевых каналов клеточной мембраны, и внеклеточный кальций поступает в цитозоль. В клетках ионы Са2+взаимодействуют с регуляторным белком кальмодулином. Комплекс кальций-кальмодулин активирует кальцийзависимые протеинкиназы, которые активируют фосфолирирование различных белков и приводят к конечным эффектам.
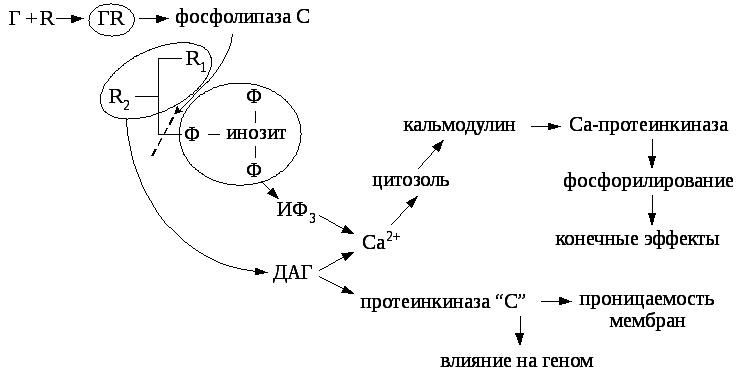
10.3.2.3. Инозитолсодержащие фосфолипиды -вторичные посредники.
Образование гормон-рецепторного комплекса активирует в клеточной мембране фосфолипазу «С», которая расщепляет фосфатидилинозит на вторичные посредники диацилглицерин (ДАГ) и инозитол-трифосфат (ИФ3). ИФ3активирует выход Са2+из внутриклеточных депо в цитозоль. Ионы кальция взаимодействуют с кальмодулином, что активирует протеинкиназы и последующее фосфолирирование белков, сопровождающееся конечными эффектами гормона. ДАГ активирует протеинкиназу «С», которая фосфорилирует сериновые или треониновые специфические белки – мишени, в результате чего может меняться проницаемость мембран, в ряде случаев через систему посредников происходит экспрессия генов.