

10.1.2. Периферійний апарат нервово-м'язової системи
Периферійний відділ нервово-м'язової системи становить опорно-руховий апарат, що дає змогу робити все розмаїття рухів людини. Однією з найважливіших частин цієї системи є м'язи, що зазнають впливу неврогенних факторів, а також ряду біологічно активних нуклеотидів (аденозинмонофосфат — АМФ), які беруть участь в обміні речовин у м'язовій тканині. Встановлено, що ослаблення або виключення дії АМФ сприяє підвищенню проникності клітинних мембран і активації в м'язових клітинах обмінних процесів.
Розвиток м'язового зусилля пов'язаний зі змінами пружності і в'язкості м'язового волокна. Напруження м'язового волокна пов'язане з його довжиною, а також зі швидкістю його подовження або напруження.
Встановлено, що один руховий нейрон іннервує тільки невелику частину м'язових волокон. При цьому між цими м'язовими волокнами розміщені інші волокна, що іннервуються від інших рухових нейронів. У фізіології і нейрофізіології прийнято називати мото- нейрон (руховий нейрон) і групу м'язових волокон, що іннервуються ним, руховою одиницею. Рухова одиниця залежно від функції м'яза містить від десятків до сотень м'язових волокон. Так, у дрібних м'язах кисті або стопи в руховій одиниці налічується кілька десятків м'язових волокон, а в двоголовому м'язі плеча, чотириголовому м'язі стегна — понад 700. Рухові одиниці в тому самому м'язі відрізняються одна від одної. Збільшення м'язової сили досягається збільшенням частоти нервових імпульсів, що надходять до рухової одиниці, або в результаті збудження нових рухових одиниць. За звичайних умов участь цих процесів є нерівнозначною і залежить від багатьох причин.
У м'язі виділено кілька типів рухових одиниць залежно від швидкості скорочення і стійкості до стомлення^ швидкі і повільні мотонейрони, що поділяються на швидкостомлювані і стійкі до втоми. Залучення в процес руху нових рухових одиниць починається з невеликих мотонейронів, що активують повільні рухові одиниці з невеликою силою, потім при збільшенні сили збудження за рахунок великих мотонейронів активуються швидкі рухові одиниці, що розвивають велику силу. Ці дані вказують на складність механізмів керування рухом, що забезпечують велику точність і різну силу рухового акту.
Будь-який рух повинен мати аферентну і еферентну системи. Без точного знання положення частин тіла в просторі і про те, як точно виконується задуманий рух, він стає неможливим. Аферентні системи — це системи висхідних провідників головного і спинного мозку, що несуть інформацію від рецепторів, які розміщені в усіх тканинах і органах, а також від усіх органів чуття. Для кожного ииду чутливості в організмі існує своя система — аналізатор. Аналізатор складається з периферійної частини — рецепторів, провідників і кіркового закінчення аналізатора із соматотопічною проекцією. В аналізаторі відбувається аналіз і синтез інформації, що надходить. Проте аналіз і синтез здійснюються також і в руховій зоні кори мозку.
Самі собою рухи поставляють організму інформацію про навколишній світ. Тільки рука й око, що рухаються, дають нам уявлення про навколишні предмети і явища. Між сенсорними і моторними системами існує дуже тісний взаємозв'язок.
Рухова функція складна і у її здійсненні важливу роль відіграють тонічні рефлекси. Навіть у розслабленому стані м'яз перебуває у певному скороченні, що й дістало назву «тонус м'яза». Так, було встановлено, що розтягування м'яза супроводжується напруженням в ньому.
Подальшими дослідження було показано, що в передніх рогах спинного мозку існують три види нервових клітин (рис. 20): великі пльфа-мотонейрони, малі мотонейрони і гамма-мотонейрони. Гам-
ма-мотонейрони становлять майже третину всіх мотонейронів передніх рогів спинного мозку.
Аксони гамма-мотонейронів і альфа-мотонейронів йдуть на периферію в передніх корінцях, а далі — у периферійних нервах.
У м'язах людини виділені більш тонкі і короткі, ніж інші, м'язові волокна. Вони знаходяться в сполучнотканинних капсулах, що мають довжину кілька міліметрів і товщину кілька десятків мікрон та форму м'язового веретена. Такі самі волокна, вміщені в капсулу, називаються інтрафузальними. На обох кінцях такого м'язового веретена розміщені тонкі волокна, що підходять до рецепторів гамма-мотонейронів. У серединній частині м'язового веретена знаходиться спіралеподібний рецептор клітини спинномозкового ганглія.
і
и
Рис.
20.
Схема
дуги міотатичного рефлексу (за Д.К.
Вогородинським, О.А. Скоромець, А.І.
ЦІваревим, 1977):
1 — альфа
мотоиейроні 2—
клітина
Реншоу; 3—
сухожилковий
рецептор Гольджі; 4
— м'язове
веретено зі спіралеподібним рецептором;
5—
гаммамотонейрон;
6—
чутливі
клітини спинномозкового ганглій; 7—
екстрапірамідальні
провідники до гамма-мотонейрона; 8—
пірамідний
шлях
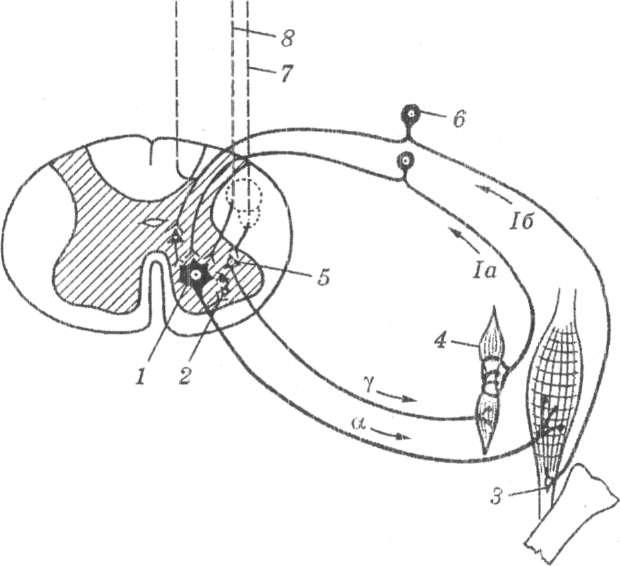
Імпульсація від гамма-мотонейронів зумовлює скорочення м'язових елементів веретена. Відбувається розтягання серединної частини м'язового веретена і виникає збудження рецепторів клітин спинномозкових гангліїв. Від спинномозкових гангліїв збудження передається на альфа-мотонейрони передніх рогів спинного мозку і виникає тонічне напруження м'яза.
Крім того, до гамма-мотонейронів передніх рогів спинного мозку надходить імпульсація від надсегментарних структур нервової системи, що беруть участь у формуванні контрактильного тонусу м'язів, а також в організації м'язових рухів (див. рис. 20). До таких надсегментальних структур належать: волокна кортико-муокулярного шляху, що бере початок від передньої центральної звивини й інших кіркових структур, ретикулярної формації стовбура мозку, підкіркових гангліїв екстрапірамідної системи, мозочка, вестибулярного ядра та інших супрасегментарних утворів.
У регуляції м'язових рухів на периферії беруть участь також нервові клітини спинномозкових гангліїв, що пов'язані з рецепторами сухожилків (сухожилкові органи Гольджі). Ці рецептори розміщені послідовно в місці переходу м'язових волокон у сухожилки і є спеціалізованими закінченнями товстих аферентних волокон. Частота розрядів сухожилкових органів Гольджі прямо залежить від м'язової сили, що розвивається. Сухожилкові рецептори мають високий поріг збудження і збуджуються тільки при виникненні значних м'язових зусиль (див. рис. 20). Збудження від цих рецепторів передається в спинний мозок, спричинюючи гальмування альфа- мотонейронів. Таке гальмування супроводжується розслабленням м'язів-синергістів і оберігає їх від надмірного перенапруження, а також приводить до одночасного скорочення м'язів-антагоністів.
У регуляції рухового акту беруть участь і мають низьку збудливість внутрішньосуглобові і зовнішньосуглобові механорецептори типу Руффіні. Вони розміщені в зв'язках і активуються при рухах тоді, коли суглоб знаходиться поблизу свого крайнього положення.
Слід зазначити, що в м'язі є багато вільних нервових закінчень, іцо також беруть участь в акті руху. Крім того, імпульсація про положення різних частин тіла надходить у кору головного мозку від рецепторів глибоких і інших видів чутливості.