

- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Электронное оглавление
- •10. Генетические функции 19
- •11. Передача информации в клетках 46
- •12. Генетический код 79
- •13. Генетический контроль синтеза днк 115
- •14. Рекомбинация 143
- •15. Регуляция экспрессии генов у прокариот 179
- •16. Регуляция экспрессии генов у эукариот 218
- •17. Генетический анализ развития 260
- •18 .Генетика соматических клеток: картирование генома человека 302
- •19. Количественные признаки 346
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая информация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •"Врожденные ошибки метаболизма" по Гэрроду
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гипотеза "один ген - один фермент"
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимические превращения при метаболизме
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены и белки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Структура белков
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутригенная комплементация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •11. Передача информации в клетках
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк
- •Синтез рнк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез белка
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Специализированный перенос информации Репликация рнк
- •Обратная транскрипция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансляция днк
- •Запрещенные (неизвестные) варианты переноса информации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (прокариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (эукариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Центральная догма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •12. Генетический код
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое изучение кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое подтверждение существования терминаторных кодонов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Расшифровка кода с помощью биохимических методов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Особенности построения генетического кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Терминаторные кодоны
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Организация нуклеотидной последовательности фагаφ χ74
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетические факторы, влияющие на трансляцию кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический код митохондрии
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •13. Генетический контроль синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полимеризация днк в репликативной вилке
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Инициация синтеза днк в точке начала репликации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Точность синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Исправление ошибок репликации и репарация днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •14. Рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Консервативный разрыв и воссоединение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Высокая отрицательная интерференция и генная конверсия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Образование структур Холлидея у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сайт-специфическая и незаконная рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Интеграция и эксцизия профагаλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Подвижные генетические элементы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая картина метаболизма днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •15. Регуляция экспрессии генов у прокариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Катаболитная репрессия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Бактериофагλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Опероны биосинтеза аминокислот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция экспрессии генов с помощью сайт-специфической рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сплайсинг гяРнк-транскриптов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Транскрипция и структура хроматина
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Согласованная регуляция экспрессии генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Метилирование днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция гемоглобиновых генов в ходе развития организма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Контроль экспрессии генов, основанный на перестройках днк
- •Типы спаривания у дрожжей
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Белки клеточной поверхности у трипаносомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Иммуноглобулиновые гены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот 243
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Заключение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •17. Генетический анализ развития
- •Дифференциальная экспрессия генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Клеточная детерминация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Эмбриогенез мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ развития
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации, затрагивающие эмбриогенез у мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации с материнским эффектом у дрозофилы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гомеозисные мутации уDrosophila
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола и дозовая компенсация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола у млекопитающих
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Механизмы генетической регуляции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •18 .Генетика соматических клеток: картирование генома человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Геном человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация клеток в культуре
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Отбор клеточных гибридов с помощью методаHat
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутрихромосомное картирование генов с помощью хромосомных перестроек
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Микроклетки и изолированные хромосомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Картирование генов с помощью днк-зондов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация in situ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая карта человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция и наследственные болезни
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Онкогены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция генов и рак
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетика соматических клеток растений
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •19. Количественные признаки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Пенетрантность и экспрессивность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены-модификаторы и гены, имеющие эпистатическое действие
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Плейотропия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Непрерывная изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Окраска семян у пшеницы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полигенное наследование
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая и средовая изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Наследуемость в различных популяциях
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Оглавление
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
17. Генетический анализ развития 257
|
| |
|
Рис. 17.8. А. Карта зачатков бластодермы D. melanogaster, построенная по принципу, описанному на рис. 17.7; показаны сайты, из клеток которых будут составлены внешние части тела имаго. Расстояния указаны в стертах. Пунктиром обозначено расстояние до средней линии мухи. Карта представлена как вид изнутри полой бластодермы. Обозначения такие же, как на рис. 17.5. Б. |
Внешние части взрослой мухи, изображенные на поверхности бластодермы. Пунктиром обозначены участки, дающие, согласно эмбриологическим исследованиям, начало нервной системе и мезодерме. (По Hotta Y, Benzer S., 1973. In : Genetic Mechanisms of Development, ed. by F. H. Ruddle, Academic Press, New York, p. 129.) |

Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
258 Экспрессия генетического материала
Карта зачатков, полученная путем детального количественного исследования гинандроморфов, обнаруживает поразительное сходство с организацией имаго по переднезадней оси (рис. 17.8). Сама яйцеклетка имеет хорошо различимые переднезаднюю и дорзально-вентральную оси, отражающие соответствующие оси самки, которая ее произвела. Построение логически согласованной карты зачатков бластодермы возможно только в том случае, если судьба тотипотентного ядра деления дробления определяется позицией, которую это ядро занимает во время формирования бластодермы. Эта ситуация является следствием случайной ориентировки первого деления дробления (рис. 17.3), на что указывает различная ориентация границы раздела полов у разных гинандроморфов.
Таким образом, в пределах оболочки яйцеклетки должна существовать некая позиционная информация, в результате реализации которой клетки, занявшие различное положение, вступают на различные пути развития.
Эмбриогенез мыши
Развитие мыши представляет интерес как тем, что может служить модельной системой для изучения развития человека, так и тем, что, сильно отличаясь от таковой у дрозофилы, она позволяет получить дополнительное представление о фундаментальных процессах развития. После оплодотворения яйцеклетки мыши зигота проходит несколько полных делений дробления; зародыш проходит стадии 2, 4, 8, 16, 32 и 64 клеток перед имплантацией в стенку матки (рис. 17.9). На стадии
|
|
|
Рис. 17.9. Ранние стадии развития в эмбриогенезе мыши. |

Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
17. Генетический анализ развития 259
|
|
|
Рис. 17.10. Карта зачатков трофобластов и внутренней клеточной массы бластоцисты мыши в возрасте 3-5 дней. (По Gardiner R. L, 1975. In: Developmental Biology of Reproduction, ed. by C. Marken and J. Papaconstantinou, Academic Press, New York.) |
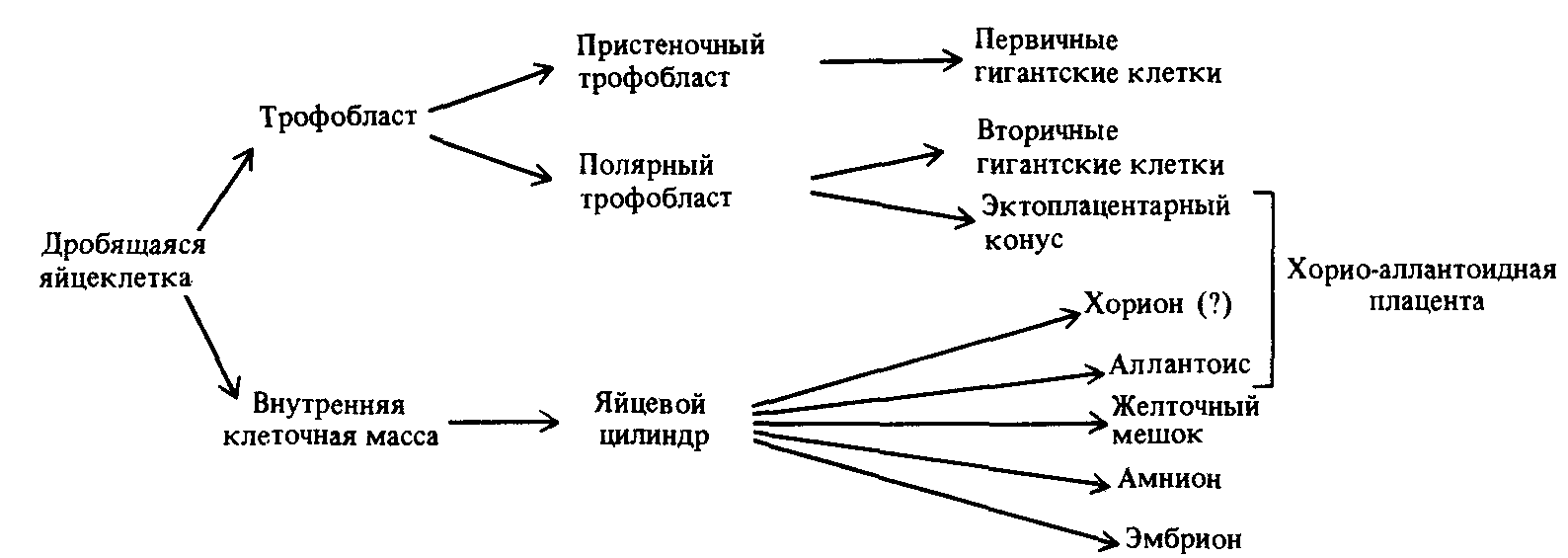
8 клеток клетки тотипотентны; каждая из них может дать начало клеткам любого типа. Тотипотентность была показана путем трансплантации клетки 8-клеточного эмбриона одного генотипа в 8-клеточный эмбрион другого генотипа и проверки генотипов клеток плаценты и зародыша, который развился из созданного таким образом эмбриона-химеры. Для определения генотипа в экспериментах часто используют электрофоретически разделяемые варианты ферментов. На стадии 64 клеток (бластоциста) эмбрион, однако, состоит уже из клеток двух хорошо различимых типов: клеток внешнего слоя, или трофобластов, и клеток внутренней клеточной массы (ВКМ). Клетки бластоцисты уже не являются тотипотентными.
Считается, что первый сигнал к тому, чтобы сестринские клетки вступили на различные пути развития, поступает на стадии 16-клеток (стадия морулы), когда в силу геометрических причин некоторые клетки оказываются со всех сторон окруженными другими клетками (inside — outside hypothesis). Вероятно, среда внутренних клеток отличается от таковой внешних клеток, которые находятся в контакте с яйцеводом. Этот позиционный сигнал вызывает первый шаг в детерминации клеток. К стадии бластоцисты эта ситуация становится очевидной благодаря разделению клеток на трофобласты и ВКМ. Однако дальнейшая судьба клеток ВКМ еще не определена. Отдельные клетки могут быть трансплантированы из ВКМ бластоцисты одного генотипа в ВКМ бластоцисты другого генотипа, где они принимают участие в процессе развития и дают начало химерным мышам, имеющим клетки двух разных генотипов. Показано, что одна клетка ВКМ дает начало дочерним клеткам, которые могут затем участвовать в образовании клеток любого типа у взрослой мыши. Сам зародыш развивается из нескольких клеток ВКМ; остальные клетки образуют ткани, не принадлежащие зародышу, такие, как плацента (рис. 17.10).