

- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Электронное оглавление
- •10. Генетические функции 19
- •11. Передача информации в клетках 46
- •12. Генетический код 79
- •13. Генетический контроль синтеза днк 115
- •14. Рекомбинация 143
- •15. Регуляция экспрессии генов у прокариот 179
- •16. Регуляция экспрессии генов у эукариот 218
- •17. Генетический анализ развития 260
- •18 .Генетика соматических клеток: картирование генома человека 302
- •19. Количественные признаки 346
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая информация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •"Врожденные ошибки метаболизма" по Гэрроду
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гипотеза "один ген - один фермент"
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимические превращения при метаболизме
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены и белки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Структура белков
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутригенная комплементация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •11. Передача информации в клетках
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк
- •Синтез рнк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез белка
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Специализированный перенос информации Репликация рнк
- •Обратная транскрипция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансляция днк
- •Запрещенные (неизвестные) варианты переноса информации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (прокариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (эукариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Центральная догма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •12. Генетический код
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое изучение кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое подтверждение существования терминаторных кодонов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Расшифровка кода с помощью биохимических методов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Особенности построения генетического кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Терминаторные кодоны
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Организация нуклеотидной последовательности фагаφ χ74
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетические факторы, влияющие на трансляцию кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический код митохондрии
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •13. Генетический контроль синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полимеризация днк в репликативной вилке
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Инициация синтеза днк в точке начала репликации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Точность синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Исправление ошибок репликации и репарация днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •14. Рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Консервативный разрыв и воссоединение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Высокая отрицательная интерференция и генная конверсия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Образование структур Холлидея у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сайт-специфическая и незаконная рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Интеграция и эксцизия профагаλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Подвижные генетические элементы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая картина метаболизма днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •15. Регуляция экспрессии генов у прокариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Катаболитная репрессия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Бактериофагλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Опероны биосинтеза аминокислот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция экспрессии генов с помощью сайт-специфической рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сплайсинг гяРнк-транскриптов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Транскрипция и структура хроматина
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Согласованная регуляция экспрессии генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Метилирование днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция гемоглобиновых генов в ходе развития организма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Контроль экспрессии генов, основанный на перестройках днк
- •Типы спаривания у дрожжей
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Белки клеточной поверхности у трипаносомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Иммуноглобулиновые гены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот 243
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Заключение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •17. Генетический анализ развития
- •Дифференциальная экспрессия генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Клеточная детерминация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Эмбриогенез мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ развития
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации, затрагивающие эмбриогенез у мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации с материнским эффектом у дрозофилы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гомеозисные мутации уDrosophila
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола и дозовая компенсация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола у млекопитающих
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Механизмы генетической регуляции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •18 .Генетика соматических клеток: картирование генома человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Геном человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация клеток в культуре
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Отбор клеточных гибридов с помощью методаHat
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутрихромосомное картирование генов с помощью хромосомных перестроек
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Микроклетки и изолированные хромосомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Картирование генов с помощью днк-зондов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация in situ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая карта человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция и наследственные болезни
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Онкогены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция генов и рак
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетика соматических клеток растений
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •19. Количественные признаки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Пенетрантность и экспрессивность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены-модификаторы и гены, имеющие эпистатическое действие
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Плейотропия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Непрерывная изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Окраска семян у пшеницы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полигенное наследование
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая и средовая изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Наследуемость в различных популяциях
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Оглавление
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
284 Экспрессия генетического материала
|
| |
|
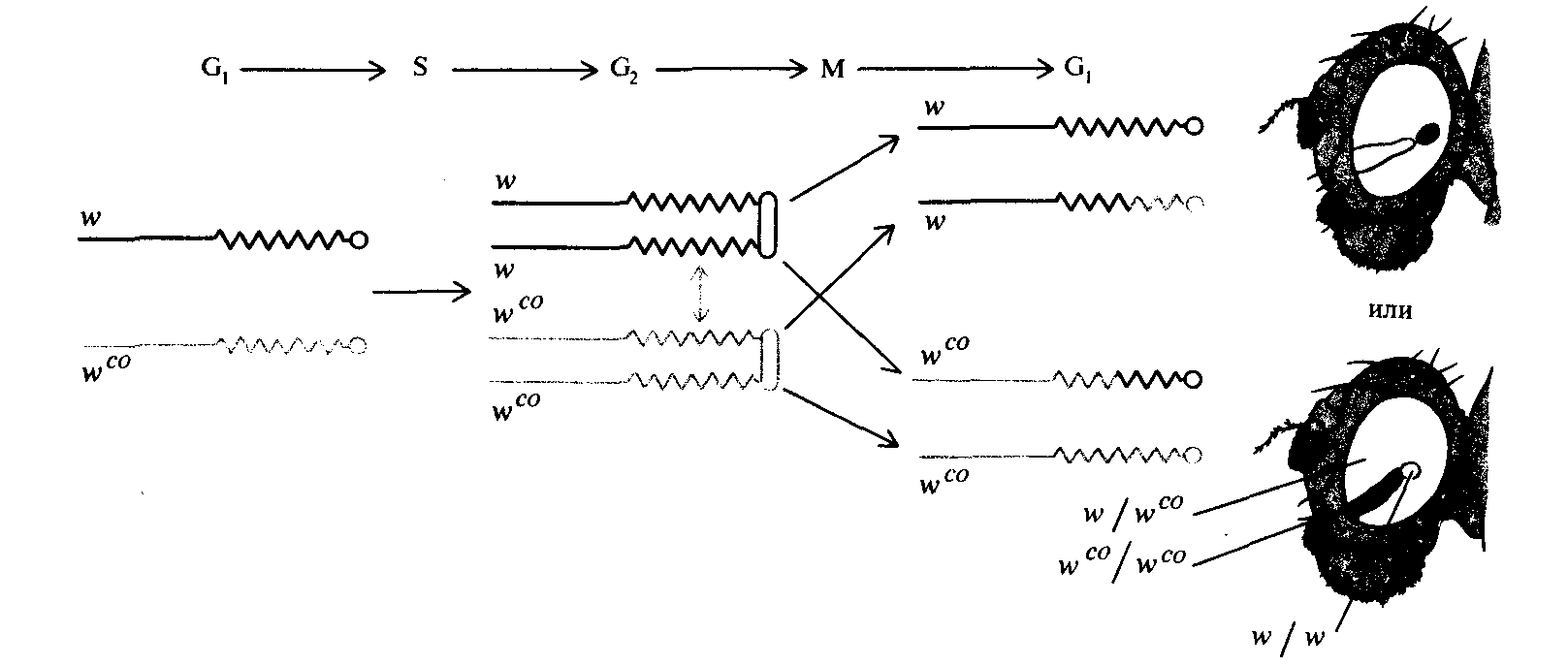
Рис. 17.25. Митотическая рекомбинация в Xхромосоме дрозофилы. Когда митотическая рекомбинация происходит в клетке глазного имагинального диска, нормальное расхождение хромосом в митозе приводит к развитию глаза с «парными пятнами». Аллели w и wco- принадлежат локусу white; глаза особи w/wco имеют темно-красный цвет. |
Заметные различия в форме и размере пятен в задней и передней части глаза свидетельствуют о влиянии положения клеток на их пролиферацию в процессе развития. (По Nöthinger R., 1972. In: Biology of Imaginai Discs, ed. by H. Ursprung, R. Nöthinger, Springer -Verlag, New York.) |
|
женных рядом пятен), каждый из которых гомозиготен по одному из этих аллелей white. Индуцированная ренгеновским облучением митотическая рекомбинация дает исследователям удобный метод изучения действия генов в процессе развития. Время облучения в процессе развития можно контролировать таким образом, что по эффекту митотической рекомбинации на признак взрослого организма определяется момент времени, когда было произведено изменение генотипа клетки. Митотическая рекомбинация, возникающая вследствие рентгеновского облучения, происходит в клетках развивающегося организма случайным образом, поэтому для того, чтобы найти клоны в интересующих исследователя органах (например, в глазах - см. рис. 17.25), необходимо проанализировать большое количество особей. Митотическая рекомбинация была использована для того, чтобы установить, требуется ли ген Рс+ только на стадии эмбриогенеза для определения экспрессии генов ВХ-С или же он необходим и на |
более поздних стадиях развития для поддержания уровня экспрессии этих генов. Эмбрионы, гомозиготные по Х-хромосоме, несущей ген yellow (у), который определяет образование желтой кутикулы и щетинок, и гетерозиготные по третьей хромосоме, имеющей в левом плече Pc + и транслоцированный ген у + , подвергаются облучению на стадии личинки (рис. 17.26). Клоны гомозиготных по Pc клеток легко идентифицировать у имаго по желтой кутикуле и щетинкам. Если такие клоны появляются на голове, тораксе или абдоминальных сегментах, их клетки дифференцируются как клетки анальной пластинки-структуры, принадлежащей эмбриональному сегменту А8. Однако при возникновении клонов в анальных пластинках их клетки дифференцируются как клетки нормальной анальной пластинки. Таким образом, удаление Pc + из клеток имагинального диска во время развития личинки приводит к активации всех генов комплекса ВХ-С, что указывает на необходимость присутствия продукта гена Pc + в имагинальных клетках для поддержания |
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
17. Генетический анализ развития 285
|
| |
|
Рис. 17.26. Зиготический генотип (слева) клетки, гетерозиготной по Polycomb + , можно изменить путем индуцированной рентгеновским облучением митотической рекомбинации, в результате чего возникает клон клеток, гомозиготных по мутантному аллелю Pc |
(справа внизу). Такой клон клеток идентифицируется у имаго по желтой кутикуле и щетинкам (фенотип yellow); эффект утраты Рс + устанавливается путем анализа типа имагинальных структур, производимых клетками клона. |

|
необходимого уровня экспрессии генов вх-с. Проведенные подобным образом эксперименты по митотической рекомбинации, в которых клоны гомозиготных по мутациям генов ВХ-С клеток получены путем облучения гетерозиготных личинок на разных стадиях их развития, показывают, что гены ВХ-С необходимы для поддержания нормального состояния клеточной детерминации напосредственно перед окукливанием. Дифференцировка клонов, индуцированных задолго до окукливания, происходит в соответствии с их мутантным генотипом. Клоны, индуциро- |
ванные незадолго до окукливания, дифференцируются нормально, что свидетельствует о присутствии во время окукливания нормальных продуктов генов BX-C, которые направляют дифференцировку по правильному пути вопреки мутантному генотипу клеток. Эти данные подтверждают гипотезу о том, что гены ВХ-С управляют процессами развития путем выбора и поддержания в клетках специфических путей развития. Выбор этих путей развития происходит в раннем эмбриогенезе в соответствии с местом расположения клеток бластодермы. |