

- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Электронное оглавление
- •10. Генетические функции 19
- •11. Передача информации в клетках 46
- •12. Генетический код 79
- •13. Генетический контроль синтеза днк 115
- •14. Рекомбинация 143
- •15. Регуляция экспрессии генов у прокариот 179
- •16. Регуляция экспрессии генов у эукариот 218
- •17. Генетический анализ развития 260
- •18 .Генетика соматических клеток: картирование генома человека 302
- •19. Количественные признаки 346
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая информация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •"Врожденные ошибки метаболизма" по Гэрроду
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гипотеза "один ген - один фермент"
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимические превращения при метаболизме
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены и белки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Структура белков
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутригенная комплементация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •11. Передача информации в клетках
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк
- •Синтез рнк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез белка
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Специализированный перенос информации Репликация рнк
- •Обратная транскрипция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансляция днк
- •Запрещенные (неизвестные) варианты переноса информации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (прокариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Колинеарность генов и полипептидов (эукариоты)
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Центральная догма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •12. Генетический код
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое изучение кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическое подтверждение существования терминаторных кодонов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Расшифровка кода с помощью биохимических методов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Особенности построения генетического кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Терминаторные кодоны
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Организация нуклеотидной последовательности фагаφ χ74
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетические факторы, влияющие на трансляцию кода
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический код митохондрии
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •13. Генетический контроль синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полимеризация днк в репликативной вилке
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Биохимический анализ репликации днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Инициация синтеза днк в точке начала репликации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Синтез днк у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Точность синтеза днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Исправление ошибок репликации и репарация днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •14. Рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Консервативный разрыв и воссоединение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Высокая отрицательная интерференция и генная конверсия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Образование структур Холлидея у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сайт-специфическая и незаконная рекомбинация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Интеграция и эксцизия профагаλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Подвижные генетические элементы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Общая картина метаболизма днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •15. Регуляция экспрессии генов у прокариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Катаболитная репрессия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Бактериофагλ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Опероны биосинтеза аминокислот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция экспрессии генов с помощью сайт-специфической рекомбинации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Участки днк, контролирующие транскрипцию
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Сплайсинг гяРнк-транскриптов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Транскрипция и структура хроматина
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Согласованная регуляция экспрессии генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Метилирование днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Регуляция гемоглобиновых генов в ходе развития организма
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Контроль экспрессии генов, основанный на перестройках днк
- •Типы спаривания у дрожжей
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Белки клеточной поверхности у трипаносомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Иммуноглобулиновые гены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •16. Регуляция экспрессии генов у эукариот 243
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Заключение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •17. Генетический анализ развития
- •Дифференциальная экспрессия генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Клеточная детерминация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Эмбриогенез мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетический анализ развития
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации, затрагивающие эмбриогенез у мыши
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Мутации с материнским эффектом у дрозофилы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гомеозисные мутации уDrosophila
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола и дозовая компенсация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Определение пола у млекопитающих
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Механизмы генетической регуляции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •18 .Генетика соматических клеток: картирование генома человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Геном человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация клеток в культуре
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Отбор клеточных гибридов с помощью методаHat
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Внутрихромосомное картирование генов с помощью хромосомных перестроек
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Микроклетки и изолированные хромосомы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Картирование генов с помощью днк-зондов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гибридизация in situ
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая карта человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция и наследственные болезни
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Онкогены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Трансфекция генов и рак
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетика соматических клеток растений
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •19. Количественные признаки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Пенетрантность и экспрессивность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Гены-модификаторы и гены, имеющие эпистатическое действие
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Плейотропия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Непрерывная изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Окраска семян у пшеницы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Полигенное наследование
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Генетическая и средовая изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Наследуемость в различных популяциях
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с. Оглавление
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
14. Рекомбинация 141
|
Рис. 14.7. Нормальные и аберрантные аски, возникающие при скрещивании мутанта с серыми спорами и штамма дикого типа (черный цвет спор) гриба Sordaria ßmicola. А. Изображены по группам обычные варианты расщепления: I. Проксимальный и дистальный варианты, образующиеся в результате расщепления при первом делении. И. Варианты чередующегося и симметричного распределения при расщеплении в ходе второго деления. Б. Нормальные и аберрантные аски распределены по группам независимо от симметрии расщепления. В нижней строчке приведены количественные данные, отражающие частоту образования каждого типа асков. (По Kitani Y. et al., 1962. Am. J. Bot, 49, 697.) |
|
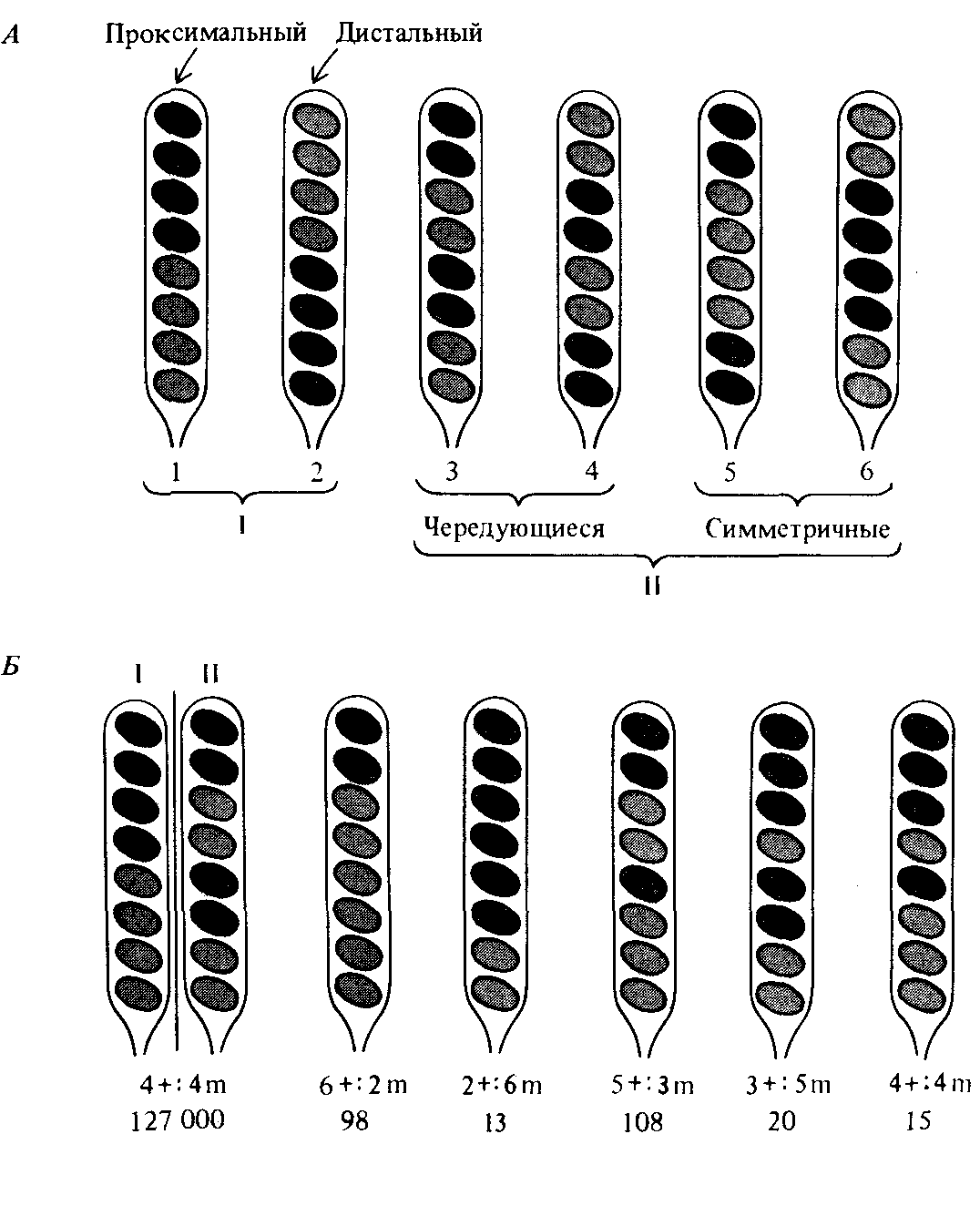
ное явление не было обнаружено и изучено у грибов и связано с другим процессом, получившим название генной конверсии. У грибов все продукты мейоза представлены в виде группы спор, расположенных внутри аска (см. гл. 5). При скрещивании двух родительских штаммов, несущих различные аллели* определенного гена, оба родительских генома оказываются в равной степени представлены в образующейся диплоидной клетке, которая впоследствии и подвергается мёйозу. Диплоидная клетка гетерозиготна - в ней присутствуют одновременно оба аллеля интересующих нас генов. Можно полагать, что образовавшиеся при мейозе и содержащиеся в одном аске споры будут характеризоваться одинаковым содержанием каждого из двух аллелей, участвовавших в скрещивании. В этом можно убедиться на примере распределения двух аллелей гена, ответственного за пигментацию спор у гриба Sordaria fimicola. Характер расщепления мутантного (серые споры) и дикого (черные споры) аллелей представлен на рис. 14.7. В зависимости от того, происходит расщепление при первом или при втором мейотическом делении, наблюдается один из вариантов расщепления, изображенных в верхней части рисунка. Каждый аск содержит по четыре черные споры дикого типа и по четыре серые мутантные споры [4 + : 4m]. Однако при анализе большого числа асков в некоторых из них обнаруживается аберрантный характер расщепления. Пять типов аберрантных асков изображены на
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 2. Пер. С англ.: – м.: Мир, 1988. – 368 с.
142 Экспрессия генетического материала
рис. 14.7 с указанием количественных данных, характеризующих частоту встречаемости для каждого типа.
A priori можно было бы полагать, что аберрантные аски возникают за счет мутаций, превращающих черные споры дикого типа в серые и, напротив, серые мутантные споры в черные в ходе мейоза. Однако внимательное изучение этого явления показало, что образование аберрантных асков не связано непосредственно с возникновением мутаций. Напротив, было установлено, что причина аберрантного расщепления заключена в явлении, связанном с рекомбинацией и получившем название генной конверсии. Генная конверсия наблюдается у многих организмов, для которых удается обнаружить все или по крайней мере половину продуктов мейотического деления. В частности, ее можно наблюдать у Drosophila благодаря использованию сцепленных Х-хромосом, позволяющему обнаружить половину образующихся тетрад.
Явление генной конверсии подробно изучалось на дрожжах Saccharomyces cerevisiae, аски которых содержат лишь четыре споры, которые в отличие от спор Sordaria или Neurospora расположены неупорядоченно. Нормальные аски, образующиеся из диплоидных дрожжевых гетерозигот, содержат аллели в соотношении 2:2, а аберрантные аски - в соотношении 3 :1 или 1 :3. Фогель, Мортимер и их коллеги провели детальное исследование конверсии гетероаллелей цистрона arg4 S. cerevisiae, кодирующего фермент аргининсукциназу (рис. 14.8). Благодаря этому исследованию можно сделать три важных заключения относительно природы процесса генной конверсии. Во-первых, примерно половина всех конверсионных явлений связана с рекомбинацией фланкирующих маркеров. Это доказывает, что конверсия не является результатом случайных мутаций, поскольку нет никаких оснований предполагать наличие взаимосвязи для процессов мутации и рекомбинации. Во-вторых, конверсия характеризуется определенной специфичностью. Гетероаллель дикого типа arg4-4 при конверсии превращается в мутантный аллель arg4-4, a дикий гетероаллель arg4-17-в мутантный arg4-17. Ясно, что случайные мутации не могут проявлять такой специфичности. В-третьих, соконеерсия (одновременная конверсия двух гетероаллелей) происходит гораздо чаще, чем это можно было бы ожидать для двух совершенно независимых событий. Это свидетель-
|
|
|
|
Рис. 14.8. Хромосома VIII Saccharomyces cerevisiae', показана тонкая структура гена arg 4. Межгенные расстояния даны в сантиморганидах, внутригенные расстояния — в нуклеотидных парах. (По Fogel S., Mortimer R., 1969. Genetics, 62, 96.) |
