

утворення
Т- і В-клітин пам'яті та підтримання їх
популяції протягом
дуже
тривалого часу: від року до десятиліть.
Стадія триває аж до на-
ступного
контакту з антигеном. Тривалість пам'яті
та її напруженість
залежать
від багатьох факторів: антигенних
властивостей збудника,
токсину
чи відповідних вакцин, анатоксинів,
інфікуючої дози, трива-
лості
контакту (перенесена хвороба, кратність
вакцинації та ін.), ста-
ну
імунної системи на час контакту з
антигеном та генетично обумо-
влених
індивідуальних особливостей імунної
відповіді. Імунологічна
пам'ять
може бути короткочасною (при поворотному
тифі, шигельоз),
тривалою
(сибірка, лептоспіроз) або навіть
пожитгєвою (поліомієліт,
кір,
коклюш). У деяких випадках пам'ять може
бути неефективною
внаслідок
швидкої мінливості збудника (грип) або
значного розмаїття
збудників
деяких захворювань (гострі кишкові
інфекції).
ня
клітин пам'яті імунна відповідь
мобілізується значно швидше і ефе-
ктивніше,
ніж при первинному контакті. Для
досягнення ефекторної
стадії
немає необхідності у тривалій стадії
індукції. Мобілізація клі-
тинної
ланки полягає у швидшому наростанні
числа антигеноспеци-
фічних
Т-кілерів. При повторній активації
гуморальних механізмів
практично
не утворюються ІдМ, антитілоутворення
розпочинається з
різкого
підвищення синтезу IgG.
Характерно,
що продукція антитіл
класу
G
розпочинається
практично відразу, а не через 1,5-2 тижні
після
контакту з антигеном (як при первинній
відповіді).
ТИПИ
ТА ОСОБЛИВОСТІ Π
РОТИ
ІНФЕКЦІЙНОГО ІМУНІТЕТУ
Визначають
3 принципові ефекторні механізми
імунного захисту:
■ >зинофільний,
В-лімфоцитарний (гуморальний) і
Т-кілерний (клітин-
ИЙ).
У
протигельмінтозному
імунітеті
домінує еозинофільна ефекторна
шка.
Т-хелпер-2
є ключовою фігурою у розвитку як алергії
негайного
ипу,
так і протипаразитарної імунної
відповіді. Клітини Лангерганса у
пкірі
і слизових оболонках вловлюють мінімальні
кількості розчинних
штигенів,
після чого мігрують у лімфатичні вузли.
Там вони перетво-
рюються
в дендритні клітини і презентують
антиген Т-хелперам-2, сто-
млюючи
їх за допомогою ІЛ-1. Т-хелпери-2 продукують
кілька типів
інтерлейкінів:
за допомогою ІЛ-4,
-13
і -16 вони стимулюють утворення
й
активність мастоцитів ("опасистих"
клітин) та ІдЕ, ІЛ-5 — активність
еозинофілів,
ІЛ-10 — пригнічують активність
Т-хелперів-1, синтез ІЛ-2
та
ІФН.4. Стадія формування та підтримки імунологічної пам'яті. Відбувається
5. Відповідь при повторному контакті з антигеном. За рахунок існуван-
Еозинофіли
належать до ефекторних клітин імунної
відповіді. Фік-
суючись
через Fc-рецептор
ІдЕ до личинок гельмінтів, вони виділяють
високоактивні
речовини. При дегрануляції еозинофілів
звільняються
катіонні
білки, пероксидаза, головний лужний
білок — вони пошко-
джують
паразита, руйнуючи його кутикулу.
Мастоцити
і базофіли, які теж активуються
Т-хелперами-2, не бе-
руть
важливої участі у пошкодженні паразитів.
Але виділяючи значну
кількість
медіаторів запалення (більше 10
інтерлейкінів, гістамін, лей-
котрієни,
фактор некрозу пухлин та ін.), ці клітини
сприяють активні-
шій
роботі еозинофілів. Надлишок біологічно
активних речовин нейт-
ралізується
за рахунок зв'язувальної функції
альбуміну, активності де-
яких
ферментів (гістаміназа, фосфоліпази)
або інгібіторів ферментів
(інгібітори
протеолізу). Ці механізми запобігають
надмірній активації
механізмів
гіперчутливості та ушкодженню тканин.
За
допомогою ІЛ-3, -4 і -6 Т-хелпери-2 стимулюють
дегрануляцію
мастоцитів
і еозинофілів. Цитокіни, що виділяються
при цьому, вто-
ринно
пришвидпгують розмноження і дозрівання
Т-хелперів-2.
В-лімфоцит
під впливом ІЛ-4 перетворюється у
плазматичну кліти-
ну
і набуває здатності синтезувати
імуноглобуліни, зокрема ІдЕ (реагі-
ни)
та IgG2a.
Реагіни
можуть відразу фіксуватися на антигенах
личин-
ки
— тоді до їх Fc-фрагмента
приєднуються еозинофіли. Можливий
варіант,
при якому ІдЕ спочатку фіксуються
Fc-фрагментами
до масто-
цитів,
а вже потім — до антигенів найпростіших.
Т-кілерний
механізм практично не задіяний у
протипаразитарному
імунітеті.
Цікаво, що Т-супресори (CDS-лімфоцити),
виділяючи ІЛ-4 та
ІЛ-10,
пригнічують активність Т-хелперів-1,
але при цьому можуть сти-
мулювати
Т-хелпери-2, продукуючи ІЛ-5 та ІЛ-13.
Гельмінти
використовують чимало механізмів, щоб
уникнути впли-
ву
імунної системи. Серед них — втрата
антигенів, наявність антиге-
нів,
спільних з антигенами господаря, сорбція
на поверхні личинки чи
дорослого
паразита антигенів господаря. Крім
того, паразити виділя-
ють
імуносупресивні речовини, а також
ініціюють реакції гіперчутли-
вості.
Останні спотворюють імунну відповідь.
Вказані особливості ство-
рюють
сприятливі умови не тільки для тривалого
виживання збудника,
але
і для розвитку інших інфекцій,
новоутворів, істинних, псевдо- і
параалергічніх
реакцій.
Протибактерійний
імунітет здебільшого
реалізується через гумо-
ральну
ланку (синтез циркулювальних антитіл)
як до самих бактерій,
так
і до їх токсинів, особливо у разі
продукції ними екзотоксину.
Спільний
вплив імуноглобулінів та системи
комплементу веде до
незворотного
пошкодження цитолеми бактерійної
клітини. Особливо
чутливою
до такого впливу комплементу є оболонка
грамнегативних
бактерій
— соматичний полісахарид у їх стінці
є потужним імуноге-
ном.
Специфічна відповідь на інкапсульовані
бактерії (клебсієла, ме-
нінгококи,
стрептококи) теж значною мірою залежить
від рівня анти-
тіл
проти антигенів клітинної мембрани.
Антитіла
можуть утворюватися не лише проти самих
бактерій, але
і
проти їх токсинів. А якщо мікроорганізм
продукує екзотоксин, то
антитоксичний
імунітет може відіграти провідну роль
у захисті органі-
зму
від інфекції (дифтерія, правець,
ботулізм). Вакцинація проти диф-
терії
чи правця теж спрямована на утворення
саме антитоксичного
імунітету.
Проте навіть достатньо напружений
антитоксичний імунітет
не
захищає людину від самого захворювання,
а лише від його усклад-
нень.
Так, успішна вакцинація проти дифтерії
лише різко знижує ймо-
вірність
розвитку міокардиту і поліневриту.
Крім
безпосередньої участі в нейтралізації
бактерій та їх токсинів,
антитіла
сприяють їх елімінації іншим чином.
Дуже важливою їх фун-
кцією
є опсонізація
—
здатність посилювати фагоцитоз бактерій.
При
цьому
антитіла взаємодіють одночасно з
антигенами бактерії (за допо-
могою
Fab-фрагмента)
і макрофага (за допомогою Fc-фрагмента).
Крім
того,
імуноглобуліни нейтралізують
антифагоцитарні фактори бакте-
рійної
стінки та речовини, які продукуються
збудником і блокують мі-
грацію
макрофагів та їх активність.
Клітинний
імунітет має важливе значення лише у
захисті від тих
бактерій,
які мають внутрішньоклітинний етап у
розвитку або є обліга-
тними
внутрішньоклітинними паразитами, як
мікобактерії, токсоплаз-
ми,
хламідії. Нейтралізація збудника
можлива тільки за тої умови, якщо
клітина,
у якій він паразитує, буде зруйнована
клітинами-кілерами.
Для
таких захворювань також характерна
гіперчутливість сповіль-
неного
типу. її наявність часто свідчить
підтвердженням активної клі-
тинної
відповіді проти даних збудників та
використовується у клінічній
практиці
для виявлення захворювання (наприклад,
туберкулінова про-
ба).
При цьому вираження шкірних проявів
при пробах, як правило,
корелює
з активністю процесу. Хоч варто зауважити,
що механізм гі-
перчутливості
не має суттєвого значення для елімінації
збудника.
Бактерійні
інфекції часто зумовлюють формування
вторинного іму-
нодефіциту,
або його поглиблення, якщо він вже
існував. Так, значне
зниження
вмісту і функціональної активності
Т-лімфоцитів описане
при
туберкульозі, лепрі, сифілісі.
Пневмо-,
гоно- і менінгококи пригнічують
активність фагоцитозу.
У
випадку їх поєднання з герпесвірусом
чи червоним пласким лишаєм
спостерігають
також пригнічення Т-ланки імунітету.
Можливо, у цьо-
му
випадку спостерігається лише посилення
розмноження персистент-
них
вірусів внаслідок супресії клітинних
механізмів, спричиненої ко-
ковою
інфекцією. Пригнічення активності
Т-лімфоцитів виявлені та-
кож
при кашлюкові, скарлатині, бруцельозі
тощо.
Специфічні
зміни в імунітеті спостерігаються
також при стрепто-
коковій
і стафілококовій інфекціях. На тлі
пригнічення Т-ланки імуні-
тету
(здебільшого Т-супресорів) спостерігається
поліклональна актива-
ція
В-лімфоцитів з можливістю синтезу
автоантитіл в осіб з генетично
детермінованою
слабкістю імуносупресорних механізмів.
Ці зміни мо-
жуть
призвести до формування ревмокардиту,
скарлатинозних уражень
серця,
ранніх постінфекційних нефритів,
артритів і васкулітів.
У
противірусному
захисті
одним з найважливіших є саме клітин-
ний
механізм захисту. Гуморальні механізми
ефективні лише після ін-
дукованої
вірусом загибелі інфікованих клітин
(це властиво для рино-,
ентеровірусів,
вірусів грипу) і виходом вірусу у
позаклітинний простір.
Проте
значна кількість вірусів позбавлена
прямої цитопатичної дії, тобто
їх
внутрішньоклітинне паразитування не
зумовлює загибелі клітини. А
деякі
— навіть стимулюють клітинний поділ
(гепаднавіруси, вірус Еп-
штейна-Барр).
Зрозуміло, що в такому випадку вірус
стає мішенню для
гуморальних
факторів лише протягом короткого
моменту переходу з
однієї
клітини в іншу. При деяких вірусних
інфекціях (герпес, цитоме-
галія)
збудник переходить з клітини в клітину
по цитоплазматичних
містках,
не руйнуючи інфікованих клітин. У таких
випадках клітинний
механізм
імунітету стає не просто важливим, а
домінуючим.
Специфічні
Т-кілери з'являються в організмі через
2-3 доби після
контакту
з вірусом, хоч їх противірусна активність
сягає максимуму
аж
наприкінці першого тижня. Т-кілери
розпізнають на поверхні інфі-
кованої
вірусом клітини комплекс "вірусний
антиген + HLA-I".
Суттєву
роль при вірусних інфекціях відіграють
також Т-ефектори
гіперчутливості
сповільненої дії. На відміну від
Т-кілерів, вони стиму-
люються
через Т-хелпери 2-го типу і реагують
переважно на антигени,
представлені
HLA-II.
Ще
одна принципова різниця полягає у
способі
знищення
інфікованих вірусом клітин. Т-кілер
здійснює його внаслі-
док
безпосереднього контакту, а Т-ефектори
гіперчутливості — на від-
стані,
виділяючи медіатори-лімфотоксини.
К-лімфоцити теж можуть
руйнувати
інфіковані вірусом клітини, але
тільки тоді, коли останні
вже
"помічені" зафіксованими на них
імуноглобулінами.
Важливе
значення у клітинній ланці захисту
мають також макрофа-
ги.
Проте вони швидше виконують роль
"прибиральників", добиваючи
клітини-мішені,
що були попередньо пошкоджені іншими
факторами
імунітету
або мічені антитілами. Інтенсивне
руйнування клітин і контакт
імунної
системи зі значною кількістю "спотворених"
антигенів, що ма-
ють
детермінанти, властиві як вірусу, так
і тканинам організму, може
призвести
до автосенсибілізації. За наявності
HLA-обумовленої
схильно-
сті
(генетично зумовлена слабкість
супресорних механізмів, що стриму-
ють
імунну відповідь) це веде до формування
автоімунних захворювань.
Імуноглобуліни
можуть зв'язувати і знешкоджувати
вірусні частин-
ки
тільки тоді, коли вони перебувають поза
клітиною. Утворені в ре-
зультаті
цього імунні комплекси досить швидко
поглинаються і пере-
травлюються
макрофагами. У деяких випадках (малі
розміри комплек-
сів,
слабкість фагоцитарної ланки, ураження
печінки та ін.) такі
комплекси
можуть тривало циркулювати в організмі
і фіксуватися на
стінках
дрібних судин з наступною активацією
комплементу і розвит-
ком
так званого імунокомплексного запалення.
На сьогодні вважаєть-
ся,
що саме персистентні та латентні вірусні
інфекції зумовлюють ура-
ження
судин і прогресування атеросклерозу.
Імуноглобуліни
можуть також зв'язуватися з вірусними
білками на
поверхні
інфікованих клітин. Інколи це призводить
до руйнування клі-
тини
внаслідок активації комплементу. Проте
частіше інфікована клі-
тина-мішень
знищується вже макрофагами або
Κ-клітинами,
які мають
рецептори
до Fc-фрагмента
імуноглобулінів. А СЗа і С5а фрагменти,
які
утворилися в результаті активації
комплементу, ініціюють запаль-
ний
процес та міграцію імуноцитів у вогнище
запалення.
Найважливішим
гуморальним фактором захисту від
вірусної інфе-
кції
є система інтерферонів, і, передусім,
γ-інтерферон.
Він продуку-
ється
Т-лімфоцитами у відповідь на вірусну
агресію і зменшує інтенси-
вність
синтезу вірусних білків в інфікованих
клітинах і вразливість до
вірусів
здорових клітин. Основними точками
впливу інтерферону є
пригнічення
транскрипції і трансляції генетичної
інформації вірусу.
Генетично
обумовлений високий рівень синтезу
інтерферону відіграє
вирішальну
роль у нечутливості деяких людей,
наприклад, до грипу.
Внаслідок
швидкої антигенної мінливості цього
вірусу, антигено-
специфічні
фактори захисту спрацьовують занадто
пізно, коли захво-
рювання
вже у розпалі. Тому саме інтерферон є
основним фактором
захисту
організму при грипі.
Протигрибковий
імунітет
забезпечується здебільшого клітинними
факторами.
Гіперчутливість сповільненого типу і
гуморальні фактори
стають
швидше свідками інфекції, ніж реальним
механізмом захисту.
Яскравим
свідченням цього є різке зростання
частоти мікозів із
віком,
коли ослабляється робота саме Т-ланки
імунітету, і принципова
неможливість
безрецидивного вилікування мікозів у
старшому віці без
супутньої
імунокорекції. Різко збільшується
ризик розвитку грибко-
вих
захворювань при патологіях, які глибоко
уражають клітинні меха-
нізми
імунної відповіді (СНІД, наркоманія,
саркоїдоз).
Лише
при глибоких мікозах формується сильна
імунна відповідь. У
випадку
поверхневих мікозів передусім реагують
механізми першої лі-
Механізми
реалізації імунної відповіді та алергії
(КЛ
- клітина Лангерганса, ДК - дендритна
клітина, МФ - макрофаг, Т-х - Т-хелпер, В
- В-
лімфоцит,
Т-к - Т-кілер, NK
- природній кілер, Κ
- кілер, Маст - мастоцит, ПК - плазматична
клітина,
Еоз - еозинофіл, АГ - антиген або алерген,
ПК - пухлинна клітина, В - клітина,
інфікована
вірусом, Γ
-
грибок, Б - бактерія).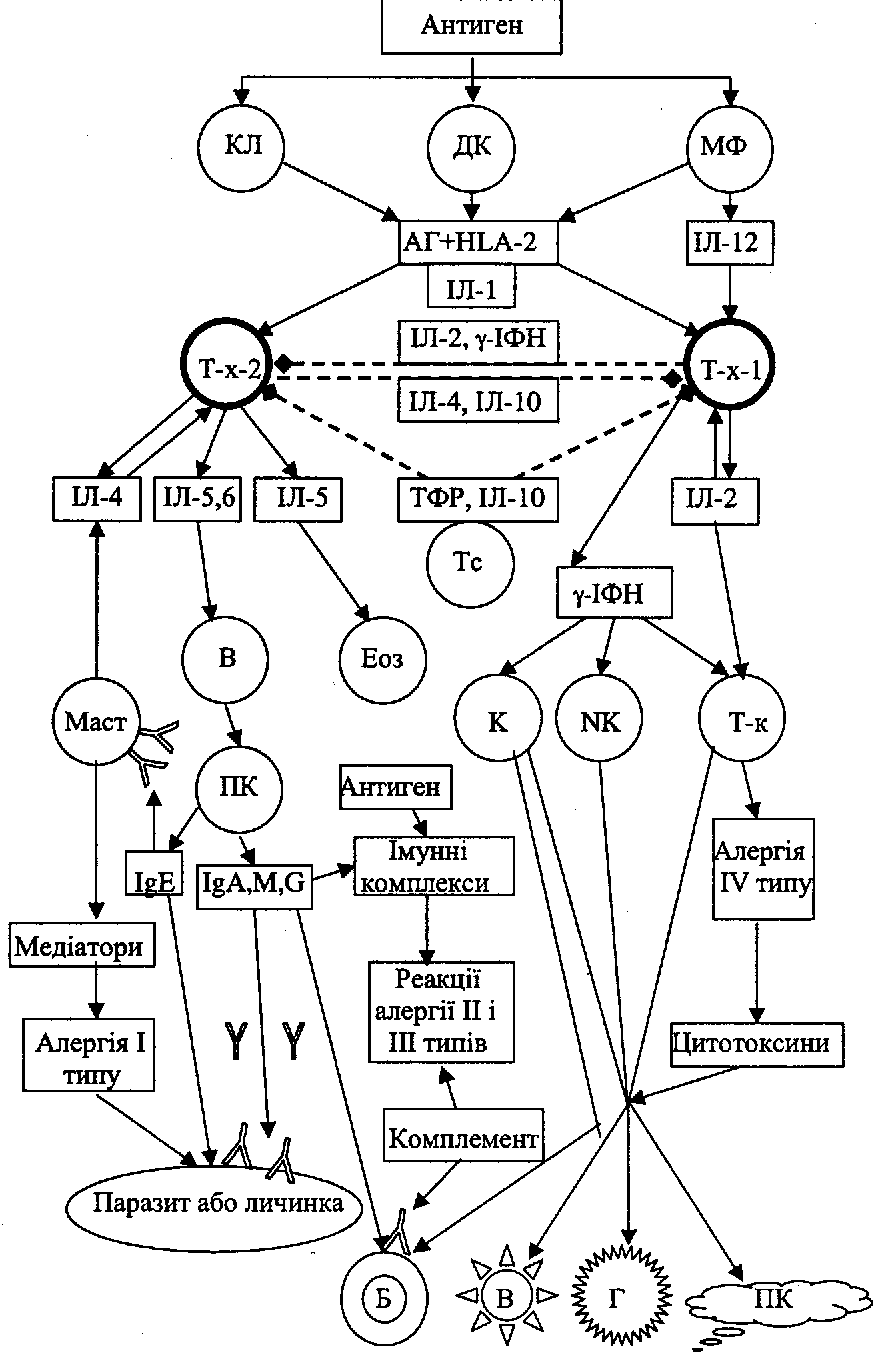
нії
захисту. їх формування поверхневих
мікозів може зумовитися навіть
факторами,
що пригнічують лише локальні механізми
імунної відповіді.
Так,
занадто тривале перебування шкіри у
неприродних умовах (вода зі
значною
концентрацією антисептиків, щільно
облягаюче взуття при си-
льній
пітливості та ін.) сприяє формуванню
стійких вогнищ поверхневої
мікотичної
інфекції — "стопи атлета", "стопи
плавця", "стопи солдата".
Протипротозоиний
Імунітет теж
забезпечується здебільшого за
рахунок
клітинних механізмів, оскільки
найпростіші належать до вну-
трішньоклітинних
збудників. Як і при мікозах, спостерігається
постій-
на
антигенна мінливість і низька напруженість
імунітету.
Наявність
лише клітинної відповіді (при слабкій
гуморальній) при-
зводить
до формування нестерильного імунітету
чи хронізації інфекції.
Ситуація
ускладнюється ще й тим, що деякі
найпростіші специфічно
пригнічують
імунну відповідь — це спостерігається
при малярії, лейш-
маніозі,
трипаносомозі, шистосомозі. Плазмодій
малярії пригнічує фу-
нкцію
макрофагів, шистосоми і трипаносоми
— надмірно посилюють
функцію
Т-супресорів. Плазмодії і трипаносоми,
крім того, зумовлю-
ють
поліклональну активацію В-лімфоцитів,
що порушує нормальний
процес
утворення антитіл.
Принципова
можливість протипріонового
імунітету на
сьогодні за-
перечується
більшістю дослідників, оскільки пріонові
білки і нормальні
білки
організму відрізняються лише за
конформацією, послідовність їх
амінокислотних
залишків та просторова орієнтація не
відрізняються.
ЗАПИТАННЯ
ДЛЯ САМОКОНТРОЛЮ
1.
Основні етапи Імунної відповіді, їх
характеристика.
2.
Особливості протигельмінтозного і
протипротозойного імунітету.
3.
Протибактерійний імунітет, механізми
його реалізації.
4.
Противірусний захист, його ефекторні
механізми.
5.
Особливості протигрибкового імунітету.
6,
Протипротозойний імунітет, механізми
його реалізації.
7.
Можливість протипріонового імунітету.
8.
Роль дисбалансу Т-хелперів 1-го і 2-го
типів при імунопатології.