

- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Электронное оглавление
- •20. Мутации генов 13
- •21. Хромосомные мутации 38
- •22. Генетическая структура популяций 78
- •23. Элементарные процессы эволюции 115
- •24. Естественный отбор 142
- •25. Инбридинг, коадаптация и географическая дифференциация 173
- •26. Видообразование и макроэволюция 208
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •20. Мутации генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Молекулярные основы генных мутаций
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Замены пар оснований
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Мутации, вызывающие сдвиг рамки считывания
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Мутагенез и репарация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Частота мутаций
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Мутагенез и эволюция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Мутации как случайный процесс
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Мутации и адаптация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •21. Хромосомные мутации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Классификация хромосомных перестроек
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Делеции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Дупликации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Инверсии
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Транслокации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Транспозиции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Робертсоновские перестройки
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Эволюция хромосом человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Анеуплоидия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Хромосомные нарушения у человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Полиплоидия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •22. Генетическая структура популяций Популяционная генетика
- •Популяции и генофонды
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Генетическая изменчивость и эволюция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Частоты генов и генотипов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Две модели популяционной структуры
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Изменчивость
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Проблема оценки генетической изменчивости
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Количественная оценка генетической изменчивости
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Полиморфизм и гетерозиготность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Электрофоретические оценки изменчивости
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Генетическая изменчивость в природных популяциях
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Полиморфизм днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •23. Элементарные процессы эволюции Эволюция - процесс двухступенчатый
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Случайное скрещивание
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Закон Харди-Вайнберга
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Применение закона Харди-Вайнберга
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Гены, сцепленные с полом
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Мутации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Миграция
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Случайный дрейф генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Эффект основателя и эффект „бутылочного горлышка"
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •24. Естественный отбор Концепция естественного отбора
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Дарвиновская приспособленность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Отбор против рецессивных гомозигот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Рецессивные летали
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Отбор против доминантных аллелей и отбор при отсутствии доминантности
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Отбор и мутации
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Оценка темпа мутирования
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Преимущество гетерозигот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Отбор против гетерозигот
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Частотно-зависимый отбор
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •25. Инбридинг, коадаптация и географическая дифференциация Коэффициент инбридинга
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Вычисление коэффициента инбридинга
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Инбредная депрессия и гетерозис
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Инбридинг в популяциях человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Генетическая коадаптация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Неравновесность по сцеплению
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Супергены
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Полиморфизм по инверсиям
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Географическая дифференциация
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Концепция расы
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Расы человека
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •26. Видообразование и макроэволюция Анагенез и кладогенез
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Концепция вида
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Процесс видообразования
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Географическое видообразование
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Квантовое видообразование
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Генетическая дифференциация в процессе видообразования
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Генетические изменения и филогения: гибридизация днк
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Филогении аминокислотных последовательностей
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Иммунология и электрофорез
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Филогении нуклеотидных последовательностей
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Теория нейтральности молекулярной эволюции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Молекулярные часы эволюции
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Эволюция структурных и регуляторных генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Эволюция размеров генома
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Эволюция посредством дупликации генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Горизонтальный перенос генов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Приложение 1. Вероятность и статистика
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •П.I. Вероятность
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •П.II. Метод хи-квадрат
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •П.III. Среднее значение и дисперсия
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •П.IV. Распределение Пуассона
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •П.V. Нормальное распределение
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Приложение 2. Ответы на задачи
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Словарь терминов
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Именной указатель
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Предметный указатель
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Указатель латинских названий
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с. Оглавление
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
- •Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
26. Видообразование и макроэволюция 247
|
| |
|
Рис. 26.26. Два строительных блока гена JgVH мыши рСh108А. Прототип А - это участок последовательности, изображенный в правой части рис. 26.25; прототип Б - на том же рисунке в центре. Под прототипами изображены их узнаваемые копии с указанием доли последовательности (в %), гомологичной прототипу. Копии расположены на |
рисунке в том же порядке, что и в гене. Числа над нуклеотидами означают их положение в полной нуклеотидной последовательности, числа внизу - положения аминокислот в полипептидной цепи. (По S. Ohno et al., 1982, Proc. Natl. Acad. Sci. USA, 79, 132-136.) |

Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
248 Эволюция генетического материала
|
| |
|
Рис. 26.27. Эволюция экзонов, кодирующих спиральный участок полипептидной цепи коллагена α 2(1) курицы. Вверху изображены нуклеотидная и кодируемая ею аминокислотная последовательности строительного блока. Неравный кроссинговер между двумя экзонами длиной по 54 п. н. мог привести к образованию экзона длиной 99 п. н. |
и остатка длиной 9 п. н. Затем в результате неравного кроссинговера между экзоном длиной 99 п. н. и одним из экзонов по 54 п. н. могли образоваться экзоны длиной 108 и 45 п.н. (По W.-H. Li. In: Evolution of Genes and Proteins, ed. by M. Nei and R. Koehn, Sinauer, Sunderland, Mass pp. 14-37, 1983.) |
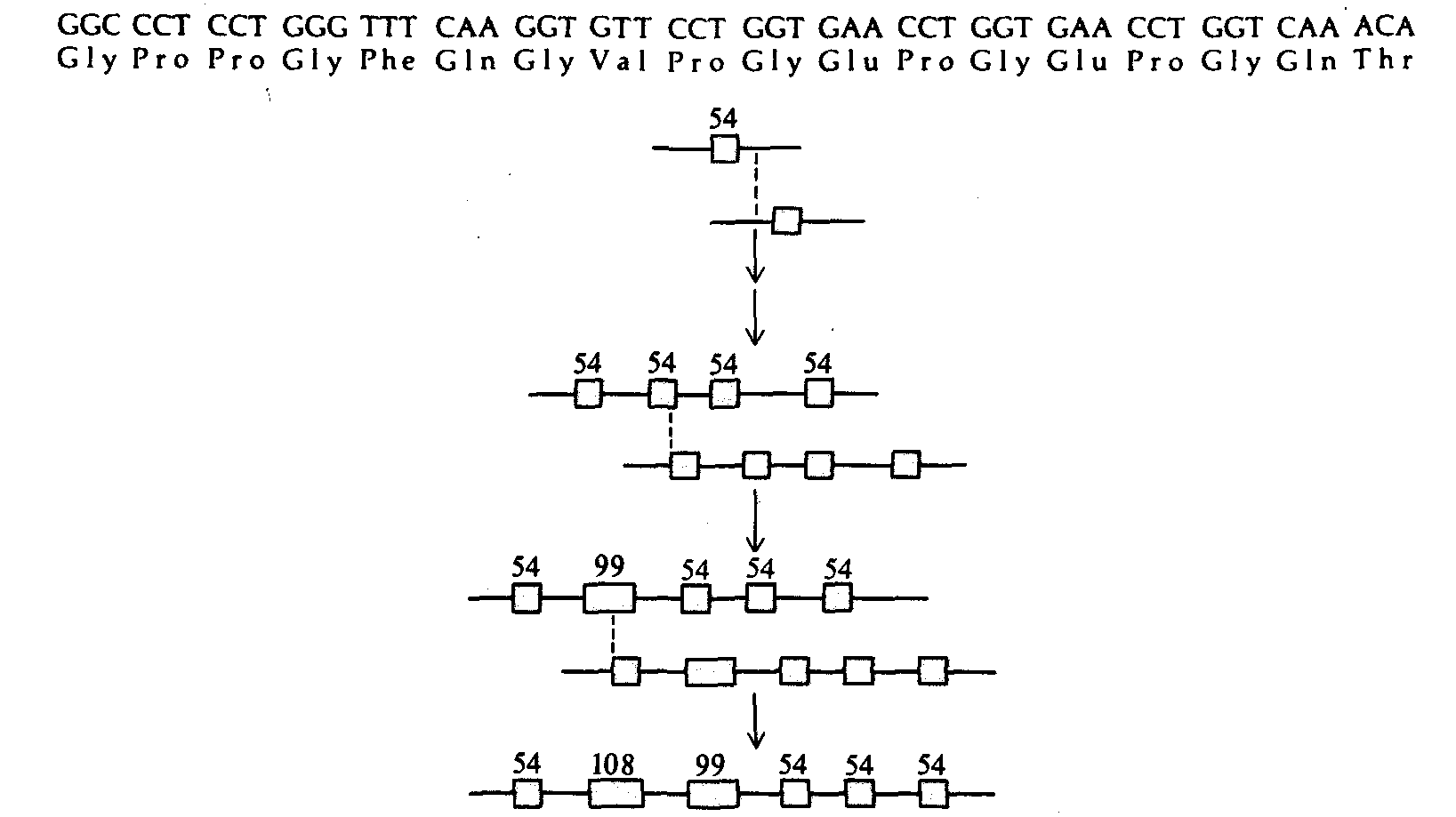
турообразующий белок костей, хрящей, соединительной ткани и кожи у позвоночных. Ген коллагена курицы имеет длину около 34 т. п. н. и содержит более 50 экзонов. Установлены нуклеотидные последовательности 21 экзона, кодирующего спираль коллагена; два из них имеют длину по 45п.н., 12 - по 54п.н., 4 - по 99 п.н. и 3 - по 108 п.н. Во всех случаях длина кратна 9 п. н. - тройке триплетов, кодирующих последовательность аминокислот типа Glu—X—Y, где X и Y часто бывают представлены пролином. На рис. 26.27 изображена гипотетическая схема эволюции этих экзонов от исходного строительного блока длиной 54 п.н. В табл. 26.11 приведены некоторые другие примеры удлинения генов посредством тандемных повторов «строительных блоков».
Некоторые гены, по-видимому, эволюционно возникли в результате слияния более мелких исходных предковых генов, осуществлявших различные функции; потомство этих предковых генов образует различные экзоны современных генов. В таких случаях каждый экзон кодирует определенный белковый домен, т.е. часть белковой молекулы с определенными функциями, гомологичными функциям, кодировавшимся простым предковым геном. Константный участок тяжелой цепи иммуноглобулина γ состоит из трех структурных доменов CH1, CH2 и СН3. Каждый домен осуществляет собственную функцию: СН3 участвует во взаимодействиях клеточной поверхности, СН2 - в фиксации комплемен-
Айала ф., Кайгер Дж. Современная генетика: в 3-х т. Т. 3. Пер. С англ.: – м.: Мир, 1988. – 336 с.
26. Видообразование и макроэволюция 249
|
Таблица 26.11. Белки с дуплицированными блоками. (По W.C. Barker et al. Atlas of Protein Sequence and Structure, vol. 5, Suppl. 3, ed by M. O. Dayhoff, National Biomédical Research Foundation, Washington, D.C., 1978, pp. 359-362.) | |||
|
Полипептид |
Общее число аминокислот |
Число аминокислот на один повтор |
Число повторов |
|
Иммуноглобулин : ε-цепь, участок С |
423 |
108 |
4 |
|
Иммуноглобулин : γ-цепь, участок С |
329 |
108 |
3 |
|
Сывороточный альбумин |
584 |
195 |
3 |
|
Парвальбумин |
108 |
39 |
2 |
|
Ингибитор протеазы сои |
71 |
28 |
2 |
|
Ингибитор протеазы подчелюстной железы |
115 |
54 |
2 |
|
Ферредоксин (Clostridium pasteurianum) |
55 |
28 |
2 |
|
Плазминоген |
790 |
79 |
5 |
|
Кальций-зависимый регуляторный белок |
148 |
74 |
2 |
|
Тропомиозин -цепи |
284 |
42 |
7 |
та, a CH1 служит точкой крепления легкой цепи. Кроме того, существует «шарнирный» участок, разделяющий две части молекулы иммуноглобулина. Рестрикционный анализ и секвенирование показывают, что ген состоит из трех экзонов, каждый из которых кодирует определенный домен белка, и четвертого экзона, кодирующего шарнирный участок.
В генах гемоглобина средний экзон кодирует домен белка, содержащий сайты, ответственные за связывание гема. Этот домен может быть предком мини-глобина, служившего переносчиком гема. Два фланговых экзона кодируют участки полипептидной цепи, окружающие продукт центрального экзона (рис. 26.28). Ген, кодирующий ADH Drosophila melanogaster, также состоит из трех экзонов, разделенных двумя интронами длиной 65 и 70 п. н. Один экзон кодирует участок из 140 аминокислотных остатков, ответственный за связь с коэнзимом. Более длинный интрон отделяет этот участок от элементов, детерминирующих каталитическую активность.
Дупликация гена часто сопровождается постепенной дивергенцией дуплицированных генов, в результате чего они приобретают в процессе эволюции различные, хотя и родственные функции. Примерами могут служить гены иммуноглобулинов (гл. 16) и глобинов (рис. 21.13). Установлено существование гомологии между дегидрогеназами, а также в других семействах ферментов, осуществляющих хотя и существенно различные, но все же родственные функции. У бактерии Acinetobacter обнаружена гомология между генами, кодирующими ферменты, которые осуществляют последовательные этапы единой цепи метаболических реакций (лактаза, декарбоксилаза, гидролаза и трансфераза); вероятно, эти гены произошли от одного предкового гена.
Псевдогенами называют участки ДНК, обладающие высокой гомологией с функциональными генами, но несущие либо nonsense-мутацию, либо мутацию со сдвигом рамки считывания, что лишает псевдогены